Т. И. Рыкова. Солеустойчивость и некоторые особенности пелагической икры и личинок белого амура и толстолобика
ВНИРО, Москва
Мы изучали особенности икры растительноядных рыб, обеспечивающие пелагический тип развития, а также отношение развивающейся икры и личинок к солености воды и содержанию в ней кислорода.
Методика опытов по выживанию икры и личинок в морской воде описана ранее (Рыкова, 1964). Интенсивность дыхания личинок определялась по методике замкнутых сосудов. Для изучения процесса образования перивителлинового пространства и прочности яйцевых оболочек пользовались методиками А. И. Зотина (1961).
Перивителлиновое пространство у рыб образуется главным образом за счет воды, поступающей под оболочку яйца из окружающей среды. Благодаря образованию перивителлинового пространства плотность яиц белого амура и толстолобика лишь немногим больше плотности воды, и икра поддерживается в пелагиали силой течения.
Мы проследили изменения плотности и объема яиц, происходящие в результате образования перивителлинового пространства с момента оплодотворения на протяжении 3-3,5 час, а в отдельных случаях - до 7 часов.
Как показали наблюдения, в первые же минуты после попадания яиц в воду отмечается резкое уменьшение их плотности и увеличение объема. Наибольшие изменения плотности яиц белого амура и толстолобика происходят в течение 40-50 мин.
Примерно таков же характер изменения объема яиц. Образование перивителлинового пространства заканчивается в основном на стадии морулы, через 80-90 мин. после оплодотворения (при температуре воды 20-23°). Через 5-10 мин. после оплодотворения заметные изменения претерпевает желток. Объем его увеличивается, из желтоватого он становится голубоватым. Эти изменения кратковременны (5-7 мин.), а затем объем желтка возвращается к исходному.
Дальнейшие изменения объема желтка связаны с начавшимся дроблением.
Таким образом, в первые минуты после оплодотворения вода поступает не только в перивителлиновую полость, но и в собственно яйцо.
Значение последующего сокращения объема желтка до исходного пока неясно.
Процесс образования перивителлинового пространства и увеличение его объема у яиц белого амура и толстолобика протекают принципиально одинаково. Объем неоплодотворенных яиц белого амура - 1,26 мм3, толстолобика - 1,07 мм3. Плотность неоплодотворенных яиц белого амура - 1,467, толстолобика - 1,424 г/см3. В результате оводнения плотность оплодотворенных яиц уменьшается до 1,015- 1,025 г/см3, а объем их увеличивается до 30-50 мм3.
При работе с икрой белого амура in vitro мы неоднократно отмечали наличие у яиц двух оболочек. Отчетливо видны они в первое время после оплодотворения, вероятно, в силу различной степени их растяжения. Позже оболочки плотно прилегают друг к другу, так что могут быть приняты за одну.
Прочность пелагических яиц, проходящих развитие в толще воды, очень мала; оболочки их тонкие и слабые.
Как показали наши наблюдения, прочность яйцевых оболочек белого амура падает к началу дробления; это снижение наблюдается и дальше, по мере увеличения числа бластомеров, и к стадии морулы величина ее становится наименьшей. Известно, (Зотин, 1961, Hayes and Armstrong, 1942), что у лососевых образование перивителлинового пространства происходит в тот промежуток времени, когда оболочки имеют сниженную прочность, и что поступление воды в яйцо прекращается с началом увеличения прочности оболочек. Мы не обнаружили повышения прочности ко времени окончания процесса образования перивителлинового пространства. Видимо, размеры перивителлинового пространства ограничивают не затвердевание оболочек.
Растворы СаСl2 и NaCl в несколько раз увеличивают прочность яйцевых оболочек белого амура. Это удалось установить, помещая оплодотворенные икринки белого амура на 5 час. в 0,1 N (децинормальные) растворы солей после предварительного выдерживания в пресной воде в течение 1-5 мин. (табл. 1).
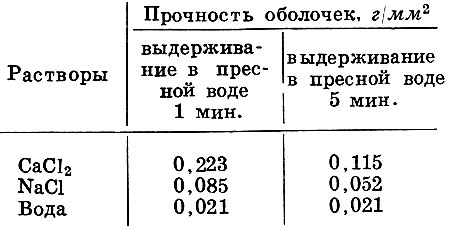
Таблица 1. Влияние солей на прочность оболочек оплодотворенных яиц белого амура
Вопрос о солеустойчивости растительноядных рыб возник в связи с вселением их в водоемы бассейнов наших южных морей и предположением возможного освоения ими приустьевых зон больших рек Азовского, Каспийского и Аральского морей, сильно зарастающих водной растительностью или богатых фитопланктоном (Карпевич, 1963). Установленные С. И. Дорошевым (1963) солевые границы выживания молоди белого амура и толстолобика принципиально допускают это предположение.
Нами получены данные по солеустойчивости икры и личинок этих рыб.
Условия инкубации контрольной, развивающейся в пресной воде, и опытной икры были одинаковыми.
Наибольшая гибель икры во время развития как в пресной воде, так и в морской отмечена в период гаструляции. Выклев же эмбрионов во всех случаях проходил почти без отхода.
Солевой диапазон выживания икры белого амура в аральской, черноморской и каспийской водах весьма сходен и находится в пределах от 0 до 5%.
При развитии икры в морской воде возникают различные отклонения от нормы, усиливающиеся с повышением солености. Значительно уменьшается объем яиц - от 46,7 мм3 в пресной воде до 0,8 мм3 в аральской воде с соленостью 14,6%. Дробление наблюдалось в опытах во всех испытуемых водах, но в аральской, приблизительно с 7%, появились неправильно дробящиеся икринки, а при 9% и выше (11,3 и 14,6%) оказывалось значительное количество недробившихся яиц. Начавшееся дробление яиц шло крайне неправильно и в большинстве случаев при солености 11,3 и 14,6% приводило к гибели икры уже на стадии морулы.
Наблюдаемые нарушения в развитии икры, такие, как "почкование" желтка и его вытекание, являются весьма характерными для осмотического действия морской воды. Развитие икры в аральской воде невысокой солености (3-5%) идет без заметных нарушений, но несколько медленнее, чем в пресной. Выклев в воде с соленостью 3% запаздывает на 3-4 часа, а с соленостью 5% - на 5-6 час. по сравнению с контролем.
У свободных эмбрионов, вылупившихся в морской воде, характерные уродства появляются при солености 3% (у одного из 19), при солености 5% - у четырех из 17.
Дальнейшее повышение солености воды приводит к еще большему числу уродов с искривленными хордами, водянкой, более слабой пигментацией глаз и тела, а также к задержке в развитии челюстного аппарата.
Подобные же нарушения в развитии наблюдались и в каспийской и черноморской водах.
Личинки в возрасте семи суток (с момента оплодотворения), помещенные в аральскую воду с соленостью 3 и 5%, за семь дней опыта не обнаружили каких-либо видимых отличий от контроля. В воде с соленостью 7,6% у некоторых личинок были разъедены солью плавники. С дальнейшим увеличением солености росло число уродливых личинок с искривленной хордой и с ярко выраженным пучеглазием. О большой степени поражения личинок солевой болезнью говорит появление при солености 9,7% язв и вздутий на плавниках и теле.
В воде с соленостью 14,6% на третьи сутки погибли все личинки.
Интенсивность дыхания личинок белого амура при помещении их непосредственно перед опытом в аральскую воду (3,7%) почти не менялась по сравнению с таковой в пресной воде, а при солености 9,7% она понижалась. Это свидетельствует об угнетенном состоянии личинок и совпадает с данными опытов по выживанию. Описанная картина прослеживалась в течение четырех суток опыта.
Полученные нами нормы потребления кислорода личинками белого амура и пестрого толстолобика (табл. 2) близки к таковым других карповых. Пороговое содержание кислорода в воде для личинок амура и толстолобика - 1,5-2 мг/л.
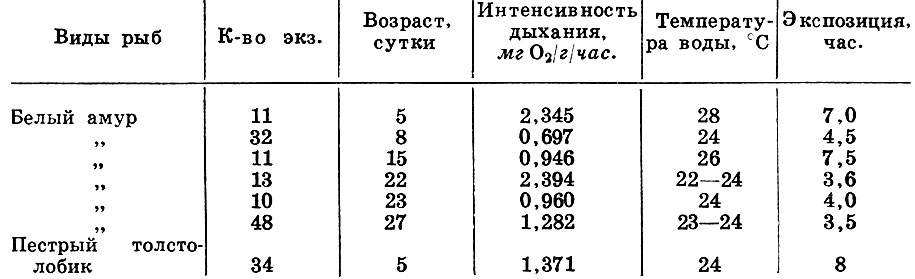
Таблица 2. Интенсивность потребления кислорода личинками белого амура, и пестрого толстолобика
1. Для икры белого амура и толстолобика характерны наличие двух оболочек, образование большого перивителлинового пространства, значительно уменьшающего плотность яиц, большая эластичность яйцевых оболочек, допускающая увеличение объема яиц в несколько десятков раз, и малая их прочность.
2. Солеустойчивость икры и свободных эмбрионов белого амура и толстолобика в морской воде лежит в пределах 0,5% и допускает успешный нерест только в пресной и слабосоленой воде.
Солеустойчивость личинок - 6-8%. Поэтому соленость воды приустьевых опресненных районов Азовского, Каспийского и Аральского морей не является фактором, препятствующим проникновению молоди в эти богатые кормом зоны.
3. По интенсивности потребления кислорода и величине кислородного порога личинки белого амура и пестрого толстолобика близки к молоди карпа.
Литература
Дорошев С. И. Выживание молоди белого амура и толстолобика в азовской и аральской воде разной солености. В кн.: "Проблемы рыбохозяйственного использования растительноядных рыб в водоемах СССР". Ашхабад, 1963.
3отин А. И. Физиология водного обмена у зародышей рыб и круглоротых. М., Изд-во АН СССР, 1961.
Карпевич А. Ф. Научные основы и перспективы акклиматизации водных организмов. В кн.: "Проблемы рыбохозяйственного использования растительноядных рыб в водоемах СССР". Ашхабад, 1963.
Рыкова Т. И. О солеустойчивости китайских растительноядных рыб на ранних стадиях развития. Труды ВНИРО, т. 55. М., 1964.
Hayes F. and Armstrong F. Physical changes in the constituent part of developing salmon eggs. Canad. J. Res., Seer. D., 20, No. 5, 1942.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'