Видовая структура рыбного населения Невской губы в весенне-летний период в связи со строительством защитной дамбы (С. В. Ружин)
УДК 639.2.053.7
Невская губа Финского залива Балтийского моря является важным рыбохозяйственным водоемом. Здесь добывается около 2 тыс. т рыбы в год, или 31% от всего улова в восточной части Финского залива (среднее по уловам за период 1976 - 1980 гг.). Промысловая рыбопродуктивность Невской губы за тот же период составила 60 кг/га.
Невская губа - естественный рыбопитомник для пресноводных и проходных рыб всей восточной части Финского залива (Широков и др., 1982; Ружин, 1984). Промысел в Невской губе и прилежащей акватории базируется на облове нерестовых скоплений рыб с весенне-летним нерестом. Вылов в апреле - июне составляет 84% от годового улова в этом районе, что указывает на наличие больших сезонных колебаний численности рыб.
Основу промысловых уловов образуют колюшка трехиглая и корюшка - 83%, на долю плотвы и ерша приходится 11%, судака и леща - 2%, на все остальные виды рыб - 4%.
Особенно возрастает значение исследования видовой структуры рыбного населения Невской губы в весенне-летний период, период максимальной численности рыб в губе, в связи со строительством комплекса сооружений для защиты Ленинграда от наводнений. Защитная дамба явится мощным антропогенным фактором, и ее строительство и функционирование могут повлиять на сложившуюся структуру рыбного населения, как это происходит обычно при реконструкции водоемов.
Исследования были проведены с целью охарактеризовать современную видовую структуру рыбного населения Невской губы и сопредельной акватории в период массовых нерестовых миграций, так как пока в литературе таких данных нет.
Материал и методика
Невская губа - крайняя восточная часть Финского залива, эстуарий Невы. На востоке ее границей служит устье Невы, на западе граница проходит по линии мыс "Лисий Нос - о. Котлин - Ломоносов. Длина губы - 21 км, наибольшая ширина 15 км, площадь водного зеркала 329 км2. Преобладающие глубины 3 - 5 м (Остов, 1971; Нежиховский, 1981). Сопредельной акваторией мы называем участки Финского залива, непосредственно примыкающие к Невской губе на западе, от Большой Ижоры и Зеленогорска до линии дамбы на востоке (см. рисунок).
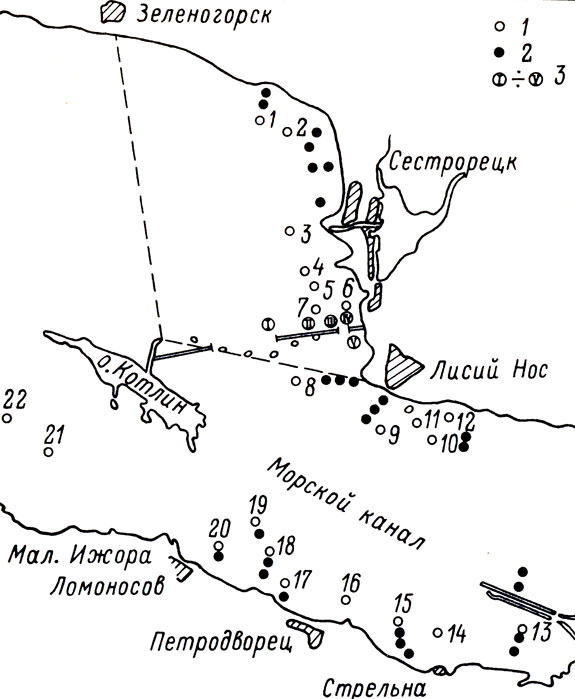
Рис. 1. Схема распределения орудий лова: 1 - орудия лова, уловы которых анализировали количественно, 2 - орудия лона, в которых учитывали только видовой состав (1984); 3 - экспериментальные орудия лова (1982)
Однако такое определение границ Невской губы, как нам кажется, следует считать устаревшим. Строительство дамбы изменило естественно-географические границы на западе. Новая граница должна проходить по телу дамбы, соответственно увеличатся протяженность и площадь района.
В основном это пресноводный район, и только в придонном слое Морского канала соленость колеблется в пределах 0,08 - 5,8‰ в течение года. Ввиду незначительных глубин, ветрового и конвекционного перемешивания преобладает гомотермия (Баранов, 1949). Граница между северной и южной частями проходит по Морскому каналу (см. рисунок).
В 1982 г. с 9 мая по 31 июля был проведен экспериментальный лов ловушками закольного типа на северном участке дамбы. Всего было выставлено 5 заколов на глубинах 1,6 - 7,2 м.
В 1984 г. анализировали уловы промысловых заколов, которые были выставлены на обычных глубинах - не более 7 м (70 проб).
Для сравнения видового состава ихтиофауны Невской губы в разные периоды использованы работы Л. С. Берга (1940) и Л. В. Широкова с соавторами (1982); в последней работе приведены результаты донных траловых съемок (1969 - 1975 гг.) в восточной части Финского залива и на одной станции в Невской губе, в районе Петродворца. Сравнивали видовые списки по показателю сходства Серенсона (Cs), эквивалентного коэффициенту Жаккара (Андреев, 1980).
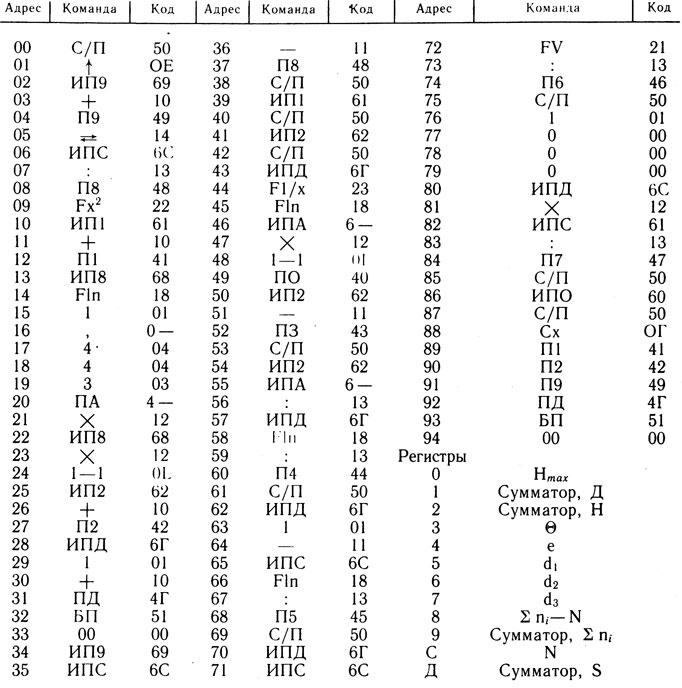
Таблица 1. Программа вычисления видовой структуры рыбного населения
Классификацию видов по критерию постоянства (C) произвели по градациям, приведенным в книге Р. Дажо (1975).

Таблица 2. Инструкция к программе (см. табл. 1)
Видовую структуру рыбного населения характеризовали показателями, взятыми из книги Ю. Одума (1975) и статьи М. М. Сметанина с соавторами (1985). Эти показатели для анализа ихтиофауны эстуариев в отечественной ихтиологической литературе использованы впервые. Для расчета величин показателей составлена и использована программа для микроЭВМ "Электроника БЗ-34" (табл. 1 и 2), записанная в общепринятых обозначениях (Цветков, Епанечников, 1984).
Вычисляются значения:
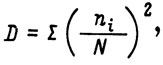
где D - индекс доминирования;
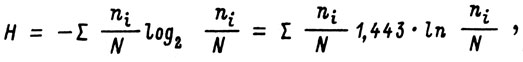
где H - показатель общего разнообразия Шеннона; ni - значимость 1-го вида (в штуках или килограммах); N - суммарная значимость всех видов (в штуках или килограммах);
где Θ - энтропийная оценка уровня абсолютной организации (ихтиоценоза); Hm=Hmax, т. е. Hm=H, когда n1=n2= ... =ni;
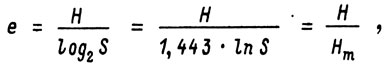
где e - показатель выровненности по Пиелу; S - число видов;
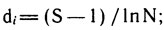
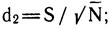
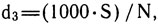
где d1, d2, d3 - показатели видового богатства или разнообразия.
Результаты исследований
Во всем исследованном районе в уловах отмечены 38 видов рыб, относящихся к 17 семействам, и 1 вид круглоротых (табл. 3). В Невской губе 35 видов из 15 семейств и минога, в сопредельной акватории - 30 видов из 14 семейств и минога.
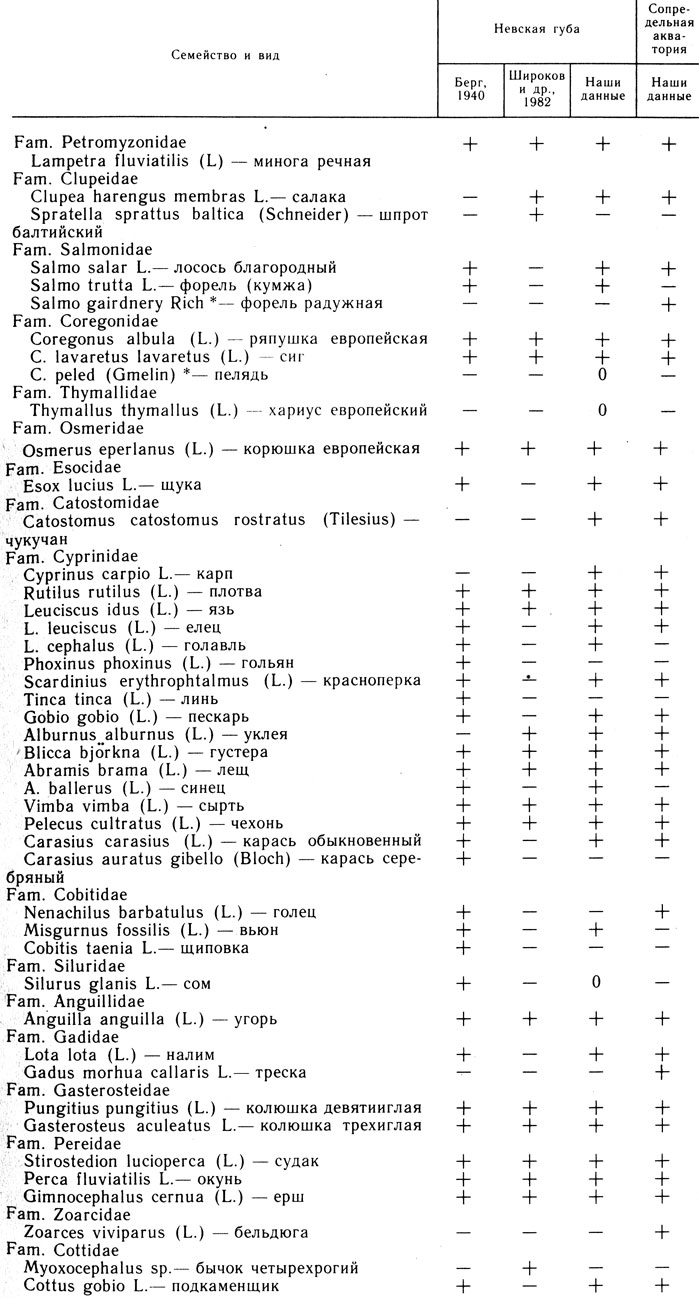
Таблица 3. Видовой состав рыбного населения Невской губы и сопредельной акватории
* (При вычислении С не учитывали, знаком "0" обозначены виды, которые в пробы 1982 и 1984 гг. не попали, но в уловах отмечены)
Среди прочих рыб обнаружены виды, прежде здесь не встречавшиеся, такие, как радужная форель, пелядь, хариус европейский; карп и чукучан уже встречались в Копорской губе Финского залива; чукучан пойман в верховьях р. Коваши (Кошелев и др., 1983) и в сопредельной Невской губе акватории в районе Северных ворот (Ружин, 1984). Хариус мог проникнуть в губу из Невы, где он и встречается (Янковская, 1955). Присутствие в исследованном районе радужной форели, пеляди, карпа и чукучана указывает на наличие процесса биологического загрязнения (Реймерс, Яблоков, 1982; Вилер, 1982). Причина этого процесса - деятельность рыбоводных предприятий. Вопрос о возможности возникновения здесь самовоспроизводящихся популяций карпа и чукучана заслуживает самостоятельного изучения.
Существует опасность проникновения в Невскую губу и сопредельную акваторию ротана (Perccottus glehni Dybowski) из водоемов Петродворца и Сестрорецка (Кудерский, 1982).
Сходство видового состава (36 видов) ихтиофаны Невской губы начала XX века (Берг, 1940) с современной ихтиофауной довольно велико: Cs=0,85 (см. табл. 3). В нашем списке отсутствуют линь, серебряный карась, голец (Nemachilus barbatulus L), щиповка - в основном бентофаги и реофилы, гольян обыкновенный, чувствительный к дефициту кислорода (Никольский, 1971).
Сходство между нашими данными и видовым списком Л. В. Широкова с соавторами (1982), насчитывающим 19 видов, меньше: Cs=0,65. В траловых уловах отсутствовали лосось, кумжа, щука, язь, голавль, красноперка, пескарь, синец, карась, вьюн, сом, налим, подкаменщик. Все это виды из группы случайных: C<25% (см. табл. 3 и 4).
Таким образом, траловые уловы позволяют объективно судить только о наличии в губе постоянных - С≥50 и добавочных видов - 50%>C≥25% (см. табл. 4). По количественному составу этих уловов, вероятно, возможна оценка колебаний относительной численности рыб или ее сезонной динамики.
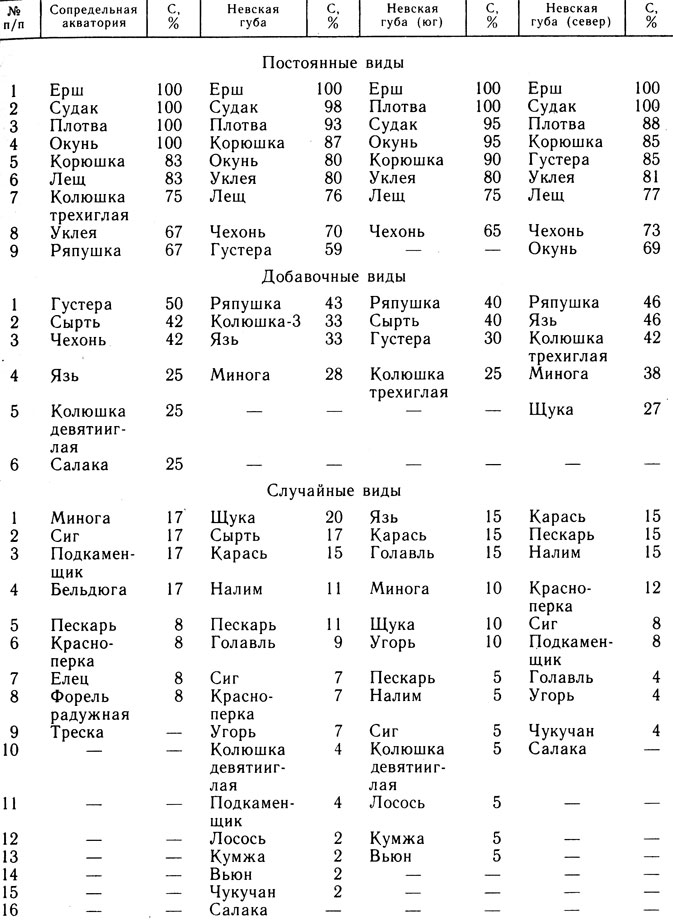
Таблица 4. Встречаемость (C) рыб Невской губы и сопредельной акватории в 1984 г.
Примечание. Знак "-" означает, что в пробу вид не попал, но в уловах встречается
Полученные данные позволили оценить сходство видового состава уловов разных участков исследованной акватории. Максимальное сходство отмечено между северным и южным участками Невской губы (Cs=0,82). На юге отсутствуют 4 случайных вида - красноперки, подкаменщик, чукучан и салака. Подкаменщик - вид оксифильный, возможно, поэтому он приурочен к зоне транзита, салака и чукучан - виды в губе очень редкие. На севере отсутствует сырть, относящаяся на юге к числу добавочных видов (C = 40%). Это можно объяснить обилием на юге водотоков - 16 против 6 на севере (Нежиховский, 1981), так как сырть проходная. Отсутствующие на севере колюшка девятииглая, вьюн, лосось, кумжа - случайные виды, вообще редки в губе.
Минимальное сходство между сопредельной акваторией и губой Cs = 0,76. Наиболее характерной чертой видового состава сопредельной акватории следует считать возрастание здесь доли представителей морского комплекса (треска, салака и бельдюга) - до 13% от общего количества видов. В Невской губе только 3%, за счет салаки, которая попадается в губе редко (C<2%). Остальные рыбы, по которым различаются эти акватории - случайные виды: щука, карась, пескарь, налим, голавль, угорь, лосось, кумжа, вьюн, елец, а также вселенцы - радужная форель и чукучан.
Существование различий в видовой структуре рыбного населения Невской губы и сопредельной акватории подтверждает и статистический анализ средних величин структурных показателей (табл. 5). Наиболее достоверны различия (p = 0,99) по показателю доминирования (D) и индексу видового разнообразия (H), рассчитанным по количеству особей. Доминирование выше в Невской губе, а видовое разнообразие - за ее пределами.
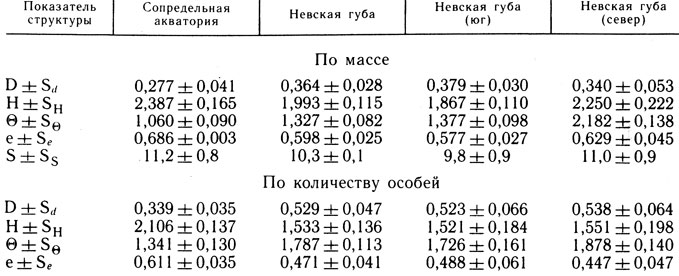
Таблица 5. Показатели структуры рыбного населения (1984 г.)
Различаются и показатели, рассчитанные по массе (p = 0,95). Уровень абсолютной организации ихтиоценоза (Θ) выше в Невской губе, а индекс видового разнообразия - за ее пределами. Несколько выше в сопредельной акватории и выровненность по Пиелу (e).
Выявленные различия в видовой структуре рыбного населения различных участков исследованной акватории могут служить характеристикой современного ее состояния. Они также могут быть использованы для анализа структурных изменений рыбного населения под воздействием различных экологических факторов, в частности, для оценки воздействий дамбы. Выявлено ядро ихтиоценоза, постоянное для всей исследованной акватории. В него входят семь видов, которые обладают постоянством не менее 50%, т. е. попадаются не менее чем в 50% уловов промысловыми орудиями лова. К ядру принадлежит прежде всего ерш (C = 100%), затем судак (98%≤С≤100%), плотва (88%≤С≤100%) и окунь (69%≤С≤100%). Остальные три вида - корюшка, уклея и лещ - не достигают 100%-ного постоянства. Корюшка к началу исследований (21 мая 1984 г.) уже в массе отнерестилась и покинула опресненные акватории. Колюшка трехиглая в основном достигла своих мелководных нерестилищ и вышла из зоны облова.
Состав ядра ихтиоценоза, так же как и другие его структурные характеристики, может меняться под воздействием изменения факторов среды (Жаков, 1984). Поэтому состав ядра также может быть использован для оценки экологических сдвигов и возможных сукцессионных процессов.
Экспериментальный лов в районе Северных ворот показал, что основу уловов составляли семь видов рыб (ерш, корюшка, плотва, окунь, лещ, судак, колюшка трехиглая). Наибольшие уловы давали ловушки III - V, расположенные на глубинах от 1,9 м до 2,7 м. Меньше уловы более глубоководных ловушек I, II (4,6 - 7,2 м). Богаче и видовой состав уловов прибрежных ловушек (III - V) - 27 видов, или 90% от общего количества выловленных видов рыб. В глубоководных ловушках (I - II) только 21 вид, или 70%.
Направление нерестовых миграций рыб с весенне-летним нерестом, составляющих основу уловов, определяется стремлением рыб достичь мелководной прибрежной зоны, где температура воды в нерестовый период выше, а соленость ниже, чем в более глубоководных районах (Ружин, 1984).
Скорости течений в водопропускных отверстиях и судоходных каналах защитной дамбы не будут превышать 0,2 - 0,3 м/с, что соизмеримо со скоростями течений в р. Неве. Поэтому дамба не является непреодолимым препятствием для миграций рыб. Однако граница распространения морских рыб станет более резкой и пройдет по телу дамбы.
В Невской губе преимущество получат более лимнофильные виды рыб, возможно, различия в структуре рыбного населения губы и сопредельной акватории станут еще более выраженными.
Наблюдения за соответствием характера промысла "Правилам рыболовства..." (1980) показали, что:
- в районе, ограниченном линией Стрельна-Морской канал - далее по Морскому каналу до Ломоносова, судак и лещ составляют от 48 до 60% от общей массы улова;
- прилов молоди судака варьировал от 67 до 100%, а леща - от 52 до 100%. Это происходит в период, когда лов судака и леща запрещен (Правила..., 1980).
Рыбоохранным организациям дана рекомендация запретить промысел рыбы в указанном районе с 20 мая по 30 июня.
Необходимо продолжить рыбохозяйственные исследования с целью разработать новый режим рыболовства в Невской губе и прилежащей акватории, так как сооружение дамбы создает условия для значительной интенсификации промысла. Пока такой режим не разработан, промысел в зоне защитной дамбы следует ограничить и вести его только под наблюдением рыбоохранных и научно-исследовательских организаций.
Андреев В. Л. 1980. Классификационные построения в экологии и систематике. М.
Баранов И. В. 1949. Годовой цикл главных гидрохимических ингредиентов воды Невской губы. - Учен. зап. ЛГУ, 126, сер. биол. наук, 21.
Берг Л. С. 1940. Рыбы Финского залива. - Изв. ВНИОРХ, 23, 2.
Вилер А. 1982. Определитель рыб морских и пресных вод Северо-Европейского бассейна. М.
Дажо Р. 1975. Основы экологии. М.
Жаков Л. А. 1984. Формирование и структура рыбного населения озер Северо-Запада СССР. М.
Кошелев Б. В., Рябов И. Н., Зимин В. А. 1984. Ихтиологические исследования Копорской губы Финского залива в 1979 - 1980 гг. - Материалы Совещания "Экологические аспекты исследования водоемов-охладителей АЭС" (март 1980 г.). М.
Кудерский Л. А. 1982. Ротан в прудах Ленинградской области. - Сб. науч. трудов ГосНИОРХ, 191.
Нежиховский Р. А. 1981. Река Нева и Невская губа. Л.
Никольский Т. В. 1971. Частная ихтиология. М.
Одум Ю. 1975. Основы экологии. М.
Остов И. М. 1971. Характерные особенности гидрологического и гидрохимического режима Финского залива как основа его рыбохозяйственного освоения. - Изв. ГосНИОРХ, 76.
Правила рыболовства во внутренних водоемах бассейна Балтийского моря. 1980.
Реймерс Н. Ф., Яблоков А. В. 1982. Словарь терминов и понятий, связанных с охраной живой природы. М.
Ружин С. В. 1984. Некоторые особенности биологии размножения рыб Невской губы и примыкающей акватории в связи со строительством дамбы для защиты Ленинграда от наводнений. - Тез. докл. III межвузовской конференции молодых ученых и специалистов, КТИРПХ. Калининград.
Сметанин М. М., Стрельников А. В., Терещенко В. Г. 1983. О применении теории шформации для анализа динамики уловов рыб в формирующихся экосистемах. - Вопр. ихтиологии, 23, 4.
Цветков А. Н., Епанечников В. А. 1984. Прикладные программы для микроЭВМ "Электроника БЗ-34", "Электроника МК-56", "Электроника МК-54". М.
Широков Л. В., Ильенкова С. А., Попов А. Н. 1982. Распределение рыб в восточной части Финского залива. - Сб. науч. трудов ГосНИОРХ, 192.
Янковская Л. А. 1955. Рыбы реки Невы и их биология и промысел. Автореф. канд. дис. Л.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'