Морские биологические ресурсы
Рост и половое созревание трески в связи с характером ее питания (Канд. биол. наук Орлова Э.Л. и Антонов С.Г., Мурманский морской биологический институт Кольского научного центра АН СССР Канд. биол. наук Ярагина Н.А., ПИНРО) (УДК 597.562-153 (268.45))
Происходящие в экосистеме Баренцева моря нарушения, связанные с почти полным выпадением из пищевой цепи звена планктофагов (мойвы, сельди, сайки), не могли не отразиться и на следующем звене - хищных рыбах, в частности на треске. Дефицит пелагических рыб в море неизбежно сказывается и на эффективности откорма трески, и на составе ее рациона, что, в свою очередь, влияет на темпы роста и полового созревания.
Нами проведены исследования этих сторон биологии трески южной части Баренцева моря в условиях изменившихся межвидовых взаимоотношений.
Материалом служили сборы по питанию и возрастному составу трески в 1986-1989 гг. Суточные рационы рассчитаны по методу Фортунатовой (1940), возраст определен по отолитам; питание исследовано у 3800 рыб, возраст - у 17769 экз.; стадии зрелости установлены визуально по 6-балльной шкале (Сорокин, 1957,1960) одновременно с определением возраста в период наиболее четких различий половозрелых и неполовозрелых рыб (ноябрь-февраль); в 1989 г. - в мае после нереста.
В рассматриваемый период колебания средней длины одновозрастных групп (табл. 1) не превышали многолетних значений, отмечаемых ранее (Пономаренко и др., 1985), и сохраняли тенденцию уменьшения разнокачественности роста с возрастом рыб. Однако в сравнении с периодами максимально высокого темпа роста (1971-1972, 1983-1985 гг.) в 1986-1989 гг. отмечены его ухудшение, значительные колебания по годам. Соответственно различались коэффициенты вариации длины: в 1986 г. при хорошем росте значения этого показателя у рыб разных возрастных групп колебались примерно на одном уровне (0,7-3,8 % у 3-5-годовиков и 0,8-2,8 % у 6-7-годовиков); в 1987 г. при ухудшении роста значения коэффициентов вариации составляли соответственно 4,4-8,1 и 2,8-4,5%, то же в 1989 г.-1,6-3,1 и 1,9-2,0% против 3,1-5,2 и 1,9-2,7% в среднем за период 1949-1978 гг. (Пономаренко и др., 1985).
Прослеживается достаточно четкая взаимосвязь темпа и характера роста трески, с условиями ее откорма в отдельные годы, сезоны. В 1986 г. интенсивный рост обеспечивался равномерным потреблением в течение 5 мес (январь - май) преимущественно мойвы и молоди сельди урожайного поколения 1983 г. (их масса составляла не менее 40-80 % массы съеденной пищи). Хотя суточные рационы в среднем не превышали 1-2 %, на долю зимне-весеннего периода пришлось более половины общего количества пищи, съеденной треской (Орлова и др., 1989). К этому же периоду (февраль - март у 4-годовиков, февраль - май у 5-7-годовиков) относится наиболее интенсивный ее рост, что отмечалось и ранее (Ярагина, 1989). Небольшое повышение приростов длины трески отмечено также в августе - сентябре после откорма креветками. В 1987 г. при чрезвычайно слабых подходах мойвы ее потребление треской наблюдалось только в отдельных районах.
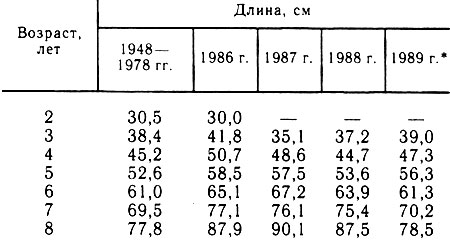
Таблица 1
* (Данные по 3-годовикам за июль, по 4-8-годовикам за сентябрь.)
В I полугодии треска распределялась в крайне западных районах (Копытовская банка) и интенсивно питалась молодью морского окуня. Нагульная миграция на восток во II полугодии началась позже обычного. Отмечены колебания величин суточных рационов трески по сезонам (0,2-3,3 % в зимне-осенний период, 0,07-2,0 % в летне-осенний). На долю зимне-весеннего периода ее откорма пришлось не более 44 % годового рациона, а роль осеннего откорма в сравнении с 1986 г., напротив, существенно возросла (с 18 до 37,5 %).
В связи с нестабильностью питания в течение сезона рост трески в возрасте до 6 лет характеризовался неравномерностью (об этом свидетельствовали высокие коэффициенты вариации длины). Только в октябре у 3-6-летних рыб появилась устойчивая тенденция к увеличению линейных приростов. Дальнейшее снижение темпа роста трески в 1988 г., по-видимому, было связано с практически полным выпадением из ее рациона мойвы и еще большим расширением пищевого спектра, хотя суточные рационы в зимний и летний периоды составляли не менее 0,8-1,0 %.
Отмечено существенное повышение роли эвфаузиид в летнем питании трески по сравнению с предыдущими годами. С 1989 г. в рационе трески все меньше становится рыбы, поэтому она переходит на ракообразных, которые являются резервным кормом. Такое питание становится регулярным и наблюдается уже с весны, но наиболее интенсивно треска потребляет ракообразных в летне-осенний период. В наибольшей степени переход на питание эвфаузиидами способствовал улучшению пищевой обеспеченности молодых рыб, в результате темп их роста по сравнению с 1988 г. увеличился.
Средняя длина рыб старшего возраста была меньше, чем в 1987 г., в связи с определенной инертностью процессов роста. Поскольку в предшествующие три года темп роста трески снижался, преодолеть отставание в росте в течение года рыбы не могли. Резкая смена качественного состава пищи сопровождалась и нарушением сезонного хода роста трески. В отличие от прошлых лет, когда она наиболее быстро росла в зимне-весенние месяцы после интенсивного откорма мойвой и молодью сельди (Ярагина, 1989), теперь ее основной рост происходил с мая по июль (линейные приросты 3-8-летних рыб составляли в среднем 5-6 см).
В целом, несмотря на существенные колебания темпа роста на протяжении четырех лет, общая картина изменилась мало в связи с низкой численностью поколений трески. Абсолютные величины годовых приростов длины находились на уровне прошлых лет, когда они составляли в среднем 7-8 см. По-видимому, в стабилизации роста трески определенную роль сыграли жировые запасы (так называемая "жирность", выражаемая в относительной массе печени). Этот показатель, достигающий 8-9 %, в 1986 г. снизился до уровня среднемноголетнего (4,1 %), а в 1987 г.-до 2,5-3,5 %. То есть темп роста и жиронакопления изменялся синхронно, но второй процесс происходил более интенсивно.
В 1989 г. при переходе трески на преимущественное питание эвфаузиидами жирность ее в летний период возросла более чем вдвое, а к концу сезона откорма - в 3 раза (до 8-9 %). Аналогичные изменения жирности при переходе с рыбно-бентосного питания на эвфаузиидное в последние годы отмечены и у другого представителя тресковых - пикши (Ковцова и др., 1989). Обычно динамику жирности связывают с пищевой обеспеченностью трески, в частности с содержанием в ее пище мойвы (Пономаренко, 1970; Ярагина, 1989, и др.).
Исследования темпа полового созревания трески проводились в контексте с основными выводами, полученными в предыдущие годы (Борисов, 1978; Пономаренко и др., 1985), о его зависимости от темпа роста рыб и структуры популяции под влиянием промысла. Последний фактор может играть решающую роль, поскольку в нерестовом стаде трески в течение многих десятилетий происходит прогрессирующее омоложение (Низовцев и др., 1988).
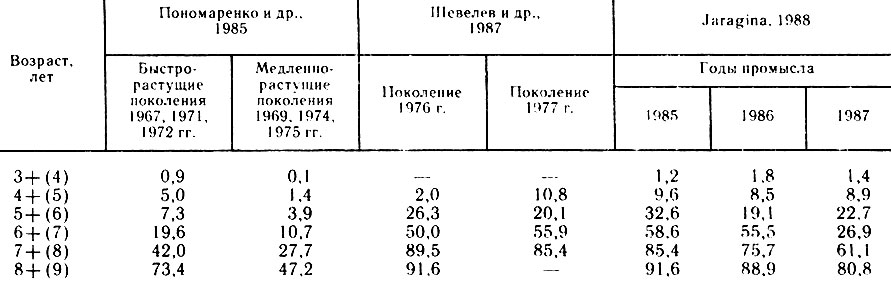
Таблица 2
Анализ данных за 80-е годы (табл. 2) свидетельствует об усилении тенденции раннего созревания трески, особенно в годы хорошего роста (1985, 1986). Однако отмечаемая в последние годы разнокачественность роста (1987 г.) в связи с ухудшением питания также может способствовать более раннему созреванию быстрорастущих рыб. Количество половозрелых рыб, наиболее лабильное в молодом возрасте, может изменяться в пределах отдельных группировок, что также, возможно, связано с их ростом. В прибрежье Мурмана в первой половине мая встречались преимущественно неполовозрелые рыбы в возрасте 3-7 лет. Во второй половине месяца отмечен подход на нагул более крупных одновозрастных рыб, среди них в большом количестве встречались отнерестовавшие 4-, 5-, и 6-годовики (соответственно 32, 33 и 81 %), которые, в свою очередь, были несколько крупнее одновозрастных неполовозрелых рыб в своей группировке.
Несмотря на отмеченные различные формы адаптаций к меняющимся условиям пищевой обеспеченности, происходящие по линии рост - половое созревание, роль раносозревающих рыб в воспроизводстве большинством исследователей признается незначительной из-за низкой плодовитости (Серебряков, 1988; Низовцев и др., 1988), а также в связи с плохой выживаемостью потомства (для трески это показано Н. В. Мухиной и Н. А. Ярагиной, 1988). Косвенным подтверждением этого является низкая численность поколений практически всех последних лет. По-видимому, современный этап формирования численности трески является критическим как с позиций обеспеченности кормом, так и возможности эффективного воспроизводства.
Основная причина сложившейся ситуации заключается в чрезмерной интенсивности промысла, оказывающего одновременно прямое воздействие на кормовую базу, численность ее основного потребителя - трески, структуру ее стада и общее состояние популяции. Положение усугубляется усилившимся в последние годы каннибализмом (Mehl, 1987; Орлова и др., 1988, Коржев, Третьяк, 1989). который в условиях низкой численности трески будет способствовать еще большему подрыву ее запасов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'