Некоторые закономерности распределения молоди рыб в районе водозабора (Е. А. Зуянов)
УДК 597 - 151:626.888
В настоящее время большое внимание уделяется экологическому способу защиты молоди рыб от попадания их в водозаборные сооружения. Этот способ предполагает изучение распределения и поведения молоди в водоемах, водотоках и зоне непосредственного влияния водозабора, с тем, чтобы прогнозировать ее распределение в районе проектируемых водозаборов, улучшить существующие конструкции рыбозащитных устройств (РЗУ) и создать новые, более эффективные.
Важность и актуальность этих исследований, недостаточность изученности данного вопроса отмечаются во многих работах по рыбозащитной тематике (Павлов, Пахоруков, 1973; Мантейфель и др., 1978; Павлов, 1979 и др.). При этом акцентируется внимание на отсутствие эффективных средств защиты рыб и большую стоимость применяемых рыбозащитных устройств.
Материал и методика. Исследования проводились в 1982 - 1983 гг. в районе действующего водозабора, расположенного на небольшом притоке Волгоградского водохранилища. Насосная станция имеет максимальную производительность 11,5 м3/с. Водозабор оснащен рыбозащитным сооружением, которое перегораживает широкий подводящий канал от водотока к насосной станции. Сбор материала по распределению молоди рыб проводили по методикам Т. С. Расса, И. И. Казановой (1966) и А. М. Пахорукова (1980). Скорости течения измеряли поплавками и гидрометрической вертушкой ГР-99. Для лова молоди использовались ихтиопланктонные ловушки ИКС-50, ИКС-80, мальковая волокуша, мальковый трал конструкции А. М. Пахорукова и А. Н. Черноусова (1982).
Определялись скорость потока на входе ловушки и трала и объемы воды, проходящей через ловушку.
За период исследований взято 684 пробы, поймано и обработано 28,2 тыс. шт. молоди рыб.
Результаты исследований. Было установлено, что выше по течению от насосной станции расположены нерестилища леща, язя, густеры, плотвы, окуня, судака, тюльки и других рыб, которые большей частью заходят на нерест из водохранилища. В мае - июле наблюдался скат ранней молоди рыб с нерестилищ.
Достигнув створа водозаборного сооружения, часть личинок продолжает движение вниз, главным образом вдоль противоположного от водозабора берега и в центральной части водотока вне зоны водозаборного течения. Другая часть попадает в это течение, где происходит накопление молоди. Концентрация личинок в 100 м3 воды в водозаборном потоке в мае - июне была выше, чем в водотоке вне этой зоны, - в среднем соответственно 33,1 и 18,2 экз., т. е. различие составило 1,8 раза.
Нами была определена граница водозаборного течения и получены усредненные значения скоростей по нескольким разрезам. Такие работы проводились параллельно со взятием ихтиологических проб. Скорость течения водозаборного потока на границе составляла 0,01 - 0,04 м/с и, постепенно увеличиваясь, около РЗУ достигала 0,35 м/с. Ширина водозаборного потока находилась в пределах 70 - 200 м.
Скорости течения на границе водозаборного потока для большинства личинок не выходят за пределы пороговых скоростей, вызывающих реореакцию. В этих условиях личинки не могут отличить водозаборное течение от транзитного в водотоке и, попав в водозаборный поток, сносятся в сторону насосной станции. Возрастающие скорости течения вызывают у личинок реореакцию. Визуальные наблюдения показали, что личинки у рыб ориентируются против течения и стараются удержаться в потоке, при этом происходит их перераспределение.
В дневное время в РЗУ сносятся более мелкие личинки рыб. Так, в мае - июне средние размеры личинок перед РЗУ были 9,3±0,4 мм, а в средней части водозаборного потока в это же время 11,4±0,9 мм. Личинки рыб, имеющие возможность сопротивляться течению (0,05 - 0,10 м/с), скапливаются днем в средней части водозаборного потока. Концентрация личинок здесь в среднем в 2,4 раза превышает концентрацию на границе водозаборного потока и в 1,7 раза - на участке непосредственно перед РЗУ (табл. 1).
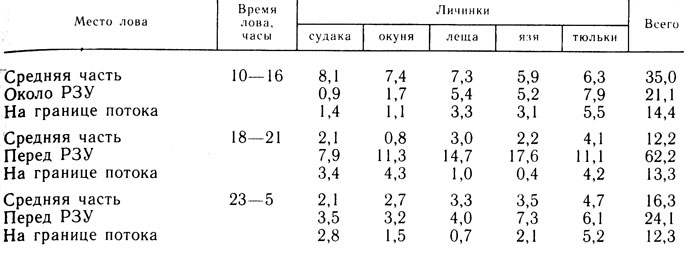
Таблица 1. Распределение личинок рыб в водозаборном потоке в течение суток за май - июнь 1982 г. (экз/100 м3)
В сумерках, в условиях ухудшения зрительной ориентации, а также, по-видимому, в связи с усталостью личинки начинают сноситься течением ближе к РЗУ, где образуют скопление, плотность которого в среднем около 62 экз/100 м3. К началу ночи (21 - 23 часа) концентрация в нем уменьшается из-за низкой эффективности РЗУ и больших скоростей течения непосредственно перед ним, превышающих критические, вследствие чего значительная часть личинок проходит за РЗУ. Ночью выраженных скоплений не образуется ни в одной зоне водозаборного потока, и концентрация личинок зависит от ската.
По отдельным видам изменения концентрации в течение суток в водозаборном потоке укладываются в приведенную схему. Исключение составляют личинки тюльки, количество которых в дневное время больше перед РЗУ. В это время личинок окуня в средней части по сравнению с участком перед РЗУ было больше в 4,4 раза, судака - в 9 раз, а леща и язя - только в 1,1 - 1,4 раза (см. табл. 1). Большее количество тюльки перед РЗУ можно объяснить ее низкой плавательной способностью, а значительная часть личинок леща и язя, скатываясь вдоль берега, просто не попадает в среднюю зону водозаборного потока, а сразу проходит и концентрируется перед РЗУ, чем в какой-то мере сглаживает различия в плотности личинок в средней зоне и перед РЗУ.
В 1983 г. на насосной станции в связи с ремонтом РЗУ был переделан водозаборный ковш. Ширина водозаборного потока уменьшилась до 12 - 50 м, в результате скорости водозаборного течения возросли в стрежневой части потока до 0,2 - 0,6 м/с. По мере удаления от стрежня скорости уменьшились до 0,01 - 0,08 м/с.
В зоне водозаборного потока также происходило перераспределение молоди рыб. Подтверждение этому - неоднократно наблюдаемый нами в экспериментальных и естественных условиях направленный выход личинок рыб размерами 9 - 20 мм в зону меньших скоростей, а также большие (в 1,8 - 3,5 раза в дневное время) концентрации личинок рыб в зоне потока со скоростями не выше 0,10 м/с по сравнению с концентрацией их в стрежневой части со скоростями более 0,10 м/с (табл. 2). Ночью, при потере зрительной ориентации, происходит уменьшение концентрации личинок на большей части аванкамеры в результате их сноса в РЗУ, а закономерно увеличивающийся в ночное время объем ската личинок рыб и отсутствие возможности выхода из потока приводит к увеличению их количества пи стрежне водозаборного потока.
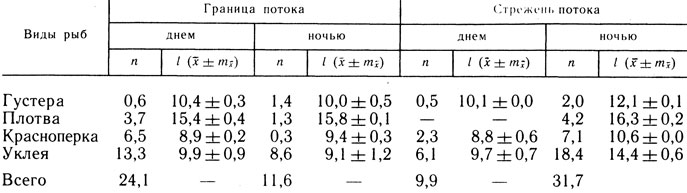
Таблица 2. Распределение личинок рыб в водозаборном потоке в течение суток в июне 1983 г.
Примечание. n - среднее количество личинок (экз/100 м3), l - средняя длина (мм).
Изменение видового состава личинок рыб в водозаборном потоке связано с тем, что в районе водозабора проводились дноуглубительные работы, что воспрепятствовало проходу на нерест леща, судака и других видов рыб, обычно нерестящихся в водотоке.
В дневное время части личинок удается выйти в "прибрежную" зону водозаборного потока и удерживаться в ней, а ночью течение выносит их сразу к РЗУ. Повышенная концентрация личинок размерами 5,9 - 18,1 мм в прибрежных частях акванкамеры (в среднем около 30 экз/100 м3) сохраняется и в ночные часы.
В течение обоих сезонов нами определялось количество проходящей за РЗУ молоди рыб. Было установлено, что в 1983 г. при относительно меньшем объеме ската в водотоке количество попадающей в водозабор молоди увеличилось в 1,6 - 2,9 раза.
Обсуждение результатов и выводы. Проведенные исследования в зоне водозаборного потока при различных гидравлических условиях показали, что в дневное время личинки рыб в зависимости от плавательной способности концентрируются в областях со скоростями течения 0,05 - 0,10 м/с, которые соответствуют крейсерским скоростям плавания личинок размерами 6 - 15 мм (Павлов, 1979). Эти скорости личинки рыб способны выдерживать часами. Данные Д. С. Павлова и Л. Г. Штафа (1981) по распределению молоди в реоградиентном потоке, полученные в экспериментальных условиях, свидетельствуют о ее стремлении находиться при освещении в зоне малых скоростей.
Указанными авторами сделан также вывод об активном перераспределении покатной молоди в темное время в зону наибольших скоростей, что в какой-то мере подтверждается нашими данными по увеличению в сумеречно-ночные часы количества более крупной молоди рыб, в стрежневой части водозаборного потока со скоростями течения 0,2 - 0,6 м/с.
Основные результаты нашей работы, определяющие распределение молоди рыб в водозаборном потоке, совпадают с положениями теоретической разработки Ю. М. Колпачкова (1981), где анализируются условия плавания и питания рыб в зоне влияния водозабора.
Мелкие, по-видимому, ослабленные личинки, а также личинки тюльки в первую очередь сносятся течением из зоны крейсерских скоростей к РЗУ. Этот процесс определяется плавательной способностью личинок и их возможностью ориентироваться в потоке (Павлов, 1979; Гирса, 1981).
Установлено, что личинки рыб, находившиеся в сумерках в прибрежных частях аванкамеры, мало подвержены влиянию водозаборного течения в ночное время.
Меньший скат личинок из прибрежий ночью, при потере зрительной ориентации, можно объяснить малыми скоростями течения у берега (0,00 - 0,04 м/с) и контактом личинок с неподвижным объектом (Гирса, 1981).
Молодь рыб, находящаяся в темное время суток в открытом потоке, не имеет возможности выйти из него из-за отсутствия зрительных и других ориентиров и сносится течением; при этом не имеет значения, естественный это водоток или канал водозабора.
Увеличение скорости водозаборного течения оказало отрицательное влияние на распределение молоди рыб, привело к большему накоплению личинок в аванкамере, и как следствие, вызвало, увеличение попадания в водозабор. Если в 1982 г. приблизительно половина личинок сносилась мимо насосной станции, то в изменившихся условиях все скатывающиеся личинки захватывались водозаборным потоком.
Снижение скорости водозаборного течения до значений, меньших крейсерских скоростей плавания личинок рыб, а также искусственное увеличение скорости транзитного потока в водотоке будут способствовать сносу личинок мимо насосной станции, меньшему накоплению и попаданию их в водозабор. При таких условиях становится возможным размещение водозаборов, аналогичных описанному на водотоках.
Гирса И. Н. 1981. Освещенность и поведение рыб. М.
Колпачков Ю. М. 1981. Условия плавания и питания молоди рыб в зоне влияния водозабора. - В кн.: Поведение рыб. М., Изд. ВАСХНИЛ.
Мантейфель Б. П., Павлов Д. С., Пахоруков А. М. 1978. Биологические основы устройства рыбозащитных и рыбопропускных сооружений.- В кн.: Биологические основы применения рыбозащитных и рыбопропускных сооружений. М.
Павлов Д. С. 1979. Биологические основы управления поведением рыб в потоке воды. М.
Павлов Д. С., Пахоруков А. М. 1973. Биологические основы защиты рыб от попадания в водозаборные сооружения. М.
Павлов Д. С., Штаф Л. Г. 1981. Распределение покатной молоди рыб в реоградиентном потоке. - ДАН СССР, 260, 2.
Пахоруков А. М. 1980. Изучение распределения молоди рыб в водохранилищах и озерах. Методическая разработка. М.
Пахоруков А. М., Черноусое А. Н. 1982. Устройство для отлова водных организмов в толще воды. А. с. 925275/СССР/ - Б. И. № 17.
Расс Т. С., Казанова И. Н. 1966. Методическое руководство по сбору личинок и мальков рыб. М.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'