Влияние голодания на вертикальное распределение молоди лососевых рыб при естественной температурной стратификации в водоеме (Л. Ю. Бугров)
УДК 597.553.2 - 151
В настоящей работе при изучении вертикального распределения рыб характер последнего рассматривался прежде всего относительно особенностей термоизбирания, свойственных молоди лососевых. Поскольку естественный температурный градиент определяется глубиной водоема и временем года, то для получения более полного и объективного материала наблюдения за поведением рыб охватывали несколько сезонов - с 1983 по 1985 г. Это дало возможность сравнить особенности распределения молоди лососевых рыб по глубине на фоне различной температурной обстановки. Такого же принципа придерживались мы и при изучении влияния состояния накормленности на поведение рыб. Так, например, молодь кижуча подвергалась воздействию голодания периодически во все сезоны (август - сентябрь, февраль, апрель, июнь - июль), а молодь атлантического лосося - только в весенне-летний период (май, июль - август).
Для изучения поведения рыб в условиях, приближенных к естественным, использовались садки-вольеры, описанные нами ранее (Бугров, 1982). Наблюдения за поведением и распределением находящихся в садках рыб проводили визуально водолазным методом с использованием подводной лаборатории "Спрут" (Муравьев, Ярвик, 1983) и дистационно посредством подводного телевидения. Для сравнения в озеро были выпущены 1000 шт. молоди кижуча того же возраста, что и в садках-вольерах, и при обнаружении рыб вне садков всякий раз регистрировалась соответствующая глубина и температура.
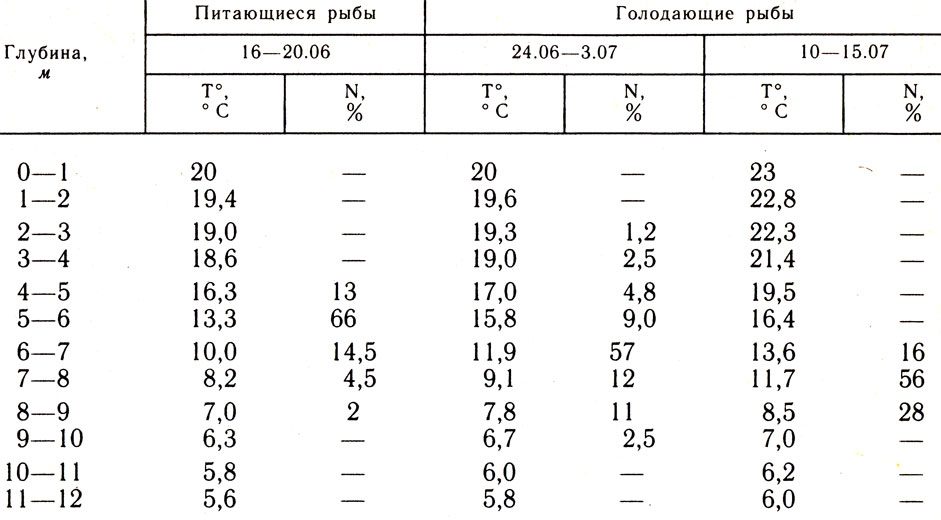
Распределение по глубине (в дневные часы) питающейся и голодающей молоди кижуча при естественной температуре стратификации
В первые дни содержания в вольерах проводилось "стартовое" голодание, когда как кижуч (в августе - сентябре), так и лосось (в мае) корма не получали в течение 20 дней. Этого периода оказалось достаточно для адаптации рыб к условиям натурного эксперимента и для установления естественного поведенческого ритма. Так, например, характерным результатом "стартового" голодания оказалось то, что оно не помешало процессу перехода подопытных рыб на уровень конечных избираемых температур (в дневные часы) и формированию суточных ритмов вертикальных миграций (Бугров, 1984).
Поскольку "стартовое" голодание протекало одновременно с адаптацией рыб, то наблюдавшийся в этот период характер распределения был результатом сложного комплекса различных факторов. Для того чтобы определить влияние именно состояния накормленности на поведение рыб, провели несколько циклов "контрольного" голодания. Трехнедельное отсутствие пищи во время зимнего и весеннего циклов достоверного воздействия на характер распределения кижуча не оказало. Единственное отличие голодавших рыб от питавшихся в эти периоды заключалось в более высокой степени реактивности первых по отношению к наблюдателю, когда присутствие аквалангиста рядом с вольером вызывало повышенную возбужденность рыб. Поэтому, во избежание искажения результатов эксперимента, наблюдения проводились из укрытия, которым служила подводная лаборатория "Спрут".
Во время жесткой температурной стратификации, которая наблюдалась на месте эксперимента (оз. Серебряное Карельский перешеек) в июле картина была иной - молодь кижуча спокойно подпускала водолаза-наблюдателя на достаточно близкую дистанцию (1,5 - 2 м). Нами представлены результаты "контрольного" голодания кижуча с 24 июня по 15 июля (см. таблицу). На первом этапе, до 3 июля, наблюдалось рассредоточение голодающих рыб на более протяженном интервале глубин по сравнению с периодом 16 - 20 июня, когда рыбы еще получали корм на фоне сходного характера температурного градиента. Если питавшаяся молодь кижуча распределялась в термодиапазоне от 16,5 до 7°С с модой в 13,3° (66% рыб), то голодающие рыбы встречались при температуре от 19,3 до 6,7°С с модальным значением избираемой температуры 11,9° (57% рыб). Интервалы глубин, на которых распределялись питавшиеся и голодавшие рыбы, составляли соответственно 4 - 8 и 2 - 9 м.
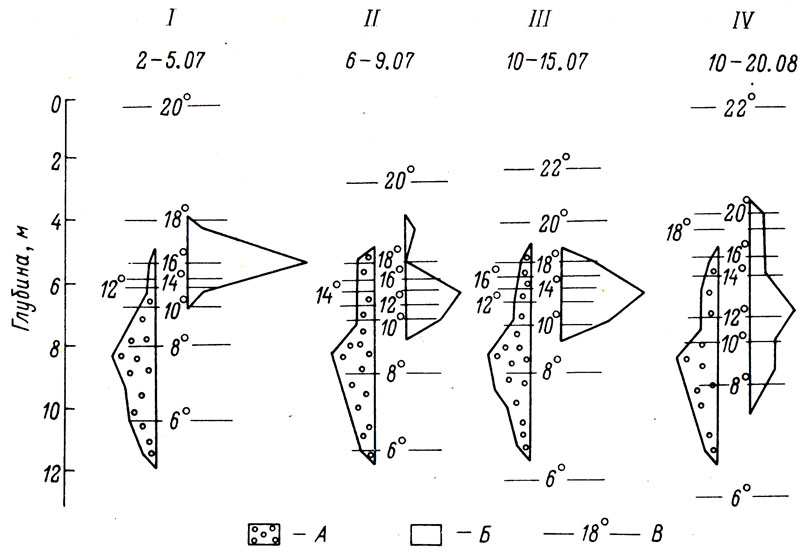
Рис. 3 Вертикальное распределение питавшейся и голодавшей молоди атлантического лосося на фоне летней температурной стратификации: I - питавшиеся рыбы; II, III, IV - голодавшие рыбы; А - ночное распределение; Б - дневное; В - изотермы
К концу периода голодания рыбы распределялись плотнее, чем на первом этапе, и даже более концентрированно, нежели питавшиеся. Модальное значение избираемой температуры осталось, однако, тем же, что и в первую декаду голодания - 11,7°, а пределы избираемой термозоны сузились до 13,6 - 8,5° и располагались в интервале глубин от 6 до 9 м (см. таблицу).
Несколько иначе реагировала на отсутствие пищи молодь атлантического лосося. Рыбы этого вида были подвергнуты голоданию продолжительностью в 6 недель, т. е. в два раза более длительному, чем молодь кижуча. В первые дни голодания, с 6 по 15 июля, было зарегистрировано распределение, аналогичное наблюдавшемуся у кижуча,- понижение модального значения избираемой температуры и расширение зоны встречаемости рыб. Если до голодания молодь лосося избирала термозону от 18 до 10° с модой 16,2°, то в первые 10 дней при отсутствии корма модальное значение избираемой температуры снизилось до 14°, а пределы температурного диапазона встречаемости рыб расширились до 19,6 - 8,8°С (см. рисунок).
В период с 10 по 20 августа, т. е. в конце цикла голодания, распределение лосося по глубине приобретает наиболее протяженный вид, охватывая интервал глубин от 3,5 до 10,5 м и температурный диапазон от 20,5 до 7°. Модальное значение избираемой температуры снижается до 12,4°, т. е. отклоняется от нормы (16,5°) на 4° (см. рисунок).
Примечательно, что в лабораторных условиях на термоградиент-приборе нам не удалось обнаружить достоверных отклонений избираемой молодью кижуча температуры при 10-дневной продолжительности опыта с голодающими рыбами (Бугров, 1984). Возможно, что это связано с необходимостью более длительного периода голодания для лабораторных условий. Однако не исключено, что причина отсутствия влияния голодания на термоизбирание рыб связана также с сезоном: молодь кижуча помещалась в термоградиентные условия зимой (Т°акклиматизации = 1,5 - 2,5°). И хотя подопытные рыбы, как уже отмечалось нами ранее (Бугров, 1982), очень быстро выходят на уровень стабильного термоизбирания (от 10 до 32 час) полной адаптации за 10-дневный срок, видимо, не происходит, поэтому и не сказывается отсутствие пищи. О необходимости учитывать сезонный фактор при изучении влияния голодания на избираемые рыбами температуры высказывались В. К. Голованов и М. И. Базаров (1981). Этими авторами сделан вывод о том, что в летний сезон фактор голодания в значительной степени влияет на термореакции молоди леща и плотвы. У леща уровень термоизбирания понижается на 4,8°, а у плотвы - на 4,7°. Для молоди окуня воздействие голодания менее значительно: избираемые температуры понижаются на 1,2° (Голованов, Базаров, 1981).
Наши результаты вполне согласуются с приведенными выше данными: за 10 суток голодания избираемая температура молоди кижуча снизилась на 1,4°, а атлантического лосося - на 2,2°С. С увеличением продолжительности срока голодания избираемая молодью лосося температура снижается еще значительней: через 35 дней на 4°.
Изменение реакций рыб на термоградиент в отсутствии пищи может быть связано с понижением уровня стандартного обмена, как это отмечалось в работах, посвященных вопросу влияния голодания на предпочитаемые температуры рыб (Дзян Яо Цин, 1959; Голованов, Базаров, 1981; Javaid, Anderson, 1967). Рассредоточение рыб в результате воздействия голодания, как нам кажется, также имеет адаптивный и, следовательно, закономерный характер - при освоении новых пространственных зон должна повышаться вероятность нахождения пищи.
Другой характерной особенностью распределения рыб на глубине, связанной с воздействием голодания, является изменение ритма вертикальных миграций. Как уже отмечалось нами ранее (Бугров, 1984, 1985), для питающейся молоди кижуча характерно обязательное наличие утреннего подъема в верхние слои воды. Вечерняя миграция выражена гораздо слабее и с возрастом ее проявление в условиях садков-вольеров и регулярного кормления постепенно затухает. Однако у голодающих рыб на четвертый день восстанавливается симметричный вид суточного ритма вертикальных миграций, т. е. отмечаются как утренний, так и вечерний пики подъема в верхние слои. Таким образом, напрашивается вывод о чисто пищевой природе стимула, вызывающего подъем молоди лососевых рыб к поверхности в сумеречное время. Определенный уровень освещенности при этом должен служить оптимальным фактором, регулирующим чередование миграционных фаз во времени суток. Следует отметить, что такое поведение молоди кижуча вполне согласуется с данными о естественной ритмике питания трехлетних рыб этого вида в природе (Зорбиди, 1977).
В свете изложенных выше факторов становится более понятным некоторое расхождение, наблюдавшееся между распределением рыб в вольерах и встречаемостью молоди кижуча вне садков. Напомним, что одновременно с зарыблением вольеров в озеро выпустили 1000 шт. молоди кижуча. Наблюдателями было зафиксировано 28 случаев встречаемости рыб этого вида около садков и других участках озера. В целом полученные в таких случаях данные оказались в границах распределения, зафиксированного в садках. В шести случаях "дикие" рыбы были встречены в утренние часы и их положение по глубине полностью совпадало с распределением кижуча в вольере, однако днем и вечером большинство из встреченных на свободе особей придерживалось несколько большей глубины. Так, например, из 12 дневных наблюдений только в трех случаях отмечалось полное совпадение с модальным значением распределения рыб в вольере, а в 9 случаях разница составляла от 1 до 3 м (различия по температуре - 1,5 - 2,5°). В вечерние часы из 8 встреч совпадение имело место также в 3 случаях, 4 рыбы были обнаружены на большей глубине (разница - 2 - 3 м), а одна рыба - под самой поверхностью. В ночные часы кижуч вне садков был встречен только 2 раза и в обоих случаях рыбы находились на расстоянии 2 - 3 м от грунта (глубина 13 - 14 м, T° = 5,7°). Реакция их на аквалангиста и освещение фонарем, так же как и в вольерах в это время суток, была сильно заторможена.
Нахождение "диких" рыб в дневные часы на более низких горизонтах, где температура на 1,5 - 2,5° ниже предпочитаемой, можно, видимо, объяснить недостаточным уровнем их обеспеченности кормом, поскольку естественная кормовая база оз. Серебряного, по имеющимся данным, отличается низкими показателями (Анцулевич, Рябова, 1982). В этих условиях рыбы вне садков испытывают перманентное голодание, которое и обусловливает специфику их вертикального распределения. Сходный же в основном характер поведения и распределения молоди кижуча в свободной воде и садках-вольерах позволяет считать, что данная методика натурного эксперимента достаточно адекватно отражает закономерности естественной ритмики вертикального распределения рыб.
На вертикальное распределение молоди лососевых рыб оказывает влияние, среди прочих факторов, также и обеспеченность пищей, причем характер распределения рыб под воздействием голодания со временем меняется и отличается следующими особенностями.
1. В контрольных условиях (при наличии корма) молодь кижуча и атлантического лосося придерживается в дневные часы слоев воды с температурой, совпадающей с конечной, избираемой в лабораторных условиях (13,6 + 0,8 и 16,5 + 0,7° - для кижуча и лосося соответственно).
2. В первые 10 дней голодающие рыбы рассредоточиваются в более протяженном интервале глубин по сравнению с питавшимися, в связи с чем увеличивается и термодиапазон встречаемости рыб. С увеличением периода голодания плотность распределения рыб периодически меняется, однако в целом наблюдается тенденция к ее уменьшению, и через шесть недель распределение приобретает наиболее протяженный вид.
3. Модальное значение избираемой рыбами температуры под воздействием голодания постепенно понижается и уже через 5 суток постоянно остается на 1,5 - 4,0° ниже, чем у питавшихся рыб.
4. Под воздействием даже непродолжительного голодания (3 - 4 дня) у трехлеток кижуча активизируется утраченная ранее вечерняя миграция в верхние слои воды, что подтверждает предположение о пищевой природе стимула, вызывающего подъем рыб в сумеречные часы.
Бугров Л. Ю. 1982. Комплексный подход к изучению преференций лососевых рыб. - Сб. науч. трудов ГосНИОРХ, 188.
Бугров Л. Ю. 1984. Особенности терморегуляционного поведения молоди лососевых рыб в лабораторных и природных условиях применительно к задачам аквакультуры. Автореф. канд. дис. Л.
Бугров Л. Ю. 1985. Особенности термопреферендума молоди кижуча при естественной температурной стратификации в водоеме. - Информ. бюл. "Биология внутр. вод", 68.
Голованов В. К., Базаров М. И. 1981. Влияние голодания на избираемые температуры у молоди леща, плотвы и окуня. - Информ. бюл. "Биол. внутр. Вод". 50.
Зорбиди Ж. К. 1977. Суточный ритм питания кижуча озера Азабачьего. - Вопр. хтиологии, 17, 1.
Муравьев В. Б., Ярвик А. Р. 1983. Опыт использования подводной пневматической камеры "Спрут" в прибрежной зоне Балтийского моря. - В кн.: Исследования по технике промышленного раболовства и поведению рыб. М., изд. ВНИРО.
Дзян Яо Цин 1959. Изменения предпочитаемых температур у некоторых видов осетровых рыб при разном уровне пищевого насыщения.- Научи, сообщ. АН СССР, 1.
Javaid M. Y., Anderson I. M. 1967. Influence of starvation on selected temperature of some salmonids. - J. Fish, Res. Board Canada, 24, 7.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'