Поведение леща в водоемах прудового типа (В. Д. Линник, Ю. В. Герасимов)
УДК 597.554.3 - 151
Проблема использования жизненного пространства животными занимает в экологии исключительно важное место (Шилов, 1977; Пианка, 1981). Исследования указанной проблемы в отношении рыб большинства внутренних водоемов затруднены ограниченной применимостью прямых наблюдений за их распределением и поведением. Большую помощь при изучении пространственных отношений у рыб оказывает биотелеметрический метод (Поддубный и др., 1979; Stasko, Pincock, 1977), посредством которого были исследованы стереотипы поведения половозрелого леща в оз. Плещееве и Рыбинском водохранилище (Малинин, Линник, 1983; Малинин и др., 1983; Линник, Герасимов, 1984). В дальнейшем для определения возможностей поведенческой адаптации нагуливающегося леща в искусственных экосистемах в 1982 г. проводили исследования на обвалованном участке прибрежья Рыбинского водохранилища и в 1983 - 1985 гг. на экспериментальных прудах Института биологии внутренних вод АН СССР.
Обвалованный участок прибрежья - водоем площадью 30 га, ограниченный земляной дамбой. Около 60% его площади зарастает жесткой водной растительностью. Водоем состоит из двух расширений - западного и восточного, соединенных каналом глубиной 1,5 м. Средняя глубина водоема 0,5 м, максимальная - 1,9 м. Восточное расширение имеет небольшую площадь, достаточные глубины, плотный грунт, бедный в качественном и количественном отношении бентос, заросли мягкой водной растительности отсутствуют. Западное расширение больше по площади глубины до 1 м, дно покрыто слоем детрита толщиной до 40 см, в середине вегетационного периода расширение на 90% зарастает мягкой водной растительностью. В бентосе обвалованного водоема доминировали хирономиды, олигохеты и моллюски. Изменения общей биомассы и агрегированности бентосных организмов представлены в табл. 1.
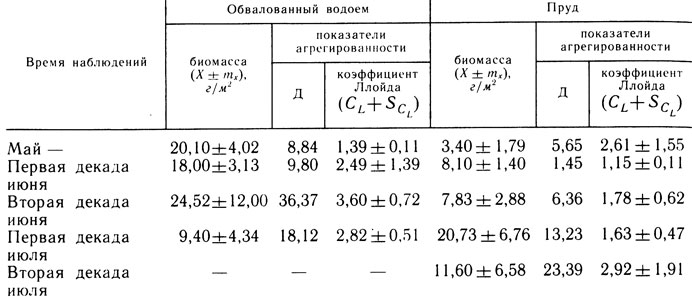
Таблица 1. Биомасса и агрегированность бентоса экспериментальных водоемов в 1982 - 1983 гг.
Примечание. Количество проб за одну съемку в обвалованном водоеме - 9 - 18, в пруду - 6.
В начале мая 1982 г. в обвалованный водоем было посажено на нерест 300 производителей леща, что при расчете на акваторию, свободную от зарослей жесткой водной растительности, дает плотность 25 шт./га.
Экспериментальные пруды - водоемы прямоугольной формы площадью 0,1 га, средней глубиной - 1,2 м, максимальной - 1,4 м. Дно прудов с небольшим уклоном к центру и водовыпуску. Зарастание мягкой и жесткой водной растительностью незначительное. Только в 1984 г. отмечено несколько большее, чем обычно, развитие мягкой растительности. Грунт - суглинок с наилком. Доминирующие группы бентоса в 1983 г.- хирономиды, моллюски и личинки ручейников, в 1984 г - моллюски и хирономиды. Общая биомасса значительно ниже, чем в 1983 г. (5,21±0,87 г/м2.), агрегированность по Ллойду (C = 1,37±0,21) статистически недостоверна (так как коэффициент Ллойда отличается от единицы менее, чем на удвоенную ошибку репрезентативности), отношение дисперсии к средней арифметической (Д) - 2,91. В 1985 г. развитие кормовой базы бентофагов было еще слабее, чем в предыдущие годы. В пруды высаживали по 30 половозрелых лещей.
Перемещение рыб прослеживали с помощью аппаратуры, разработанной ВНИРО (Саранчов, 1984). Ультразвуковые передатчики АРМ-30-15 диаметром 17 мм, длиной 35 мм, весом в воде 5,5 г и сроком действия более 40 суток крепили наружно у основания спинного плавника рыб (Gray, Haynes, 1979), предварительно анестезированных хинальдином. Всего помечено 37 особей с длиной тела 37,2 - 43,2 см. В 1982 - 1983 гг. лещей метили в первой - второй декадах мая, в 1984 г. - во второй декаде июня, в 1985 г. - в первой декаде июня. Срок наблюдений - 25 - 48 суток.
Во время периодически проводимых суточных станций каждые 10 - 15 мин регистрировали местоположение меченых особей. Использовали метод триангуляции (Поддубный и др., 1979), дающий координаты меченых рыб с ошибкой не более ±2 м. По трассам перемещений лещей вычисляли скорость плавания, неравномерность посещаемости участков водоема - по коэффициенту Ллойда и информационному показателю относительной организации системы R Ферстера (Антомонов, 1977). Учитывая особенности варьирования скоростей плавания рыб (сильную положительную асимметричность и перерассеянность), в качестве корректного показателя использовали среднюю геометрическую (Mackay, Craig, 1983).
В обвалованном водоеме в начале периода наблюдений для рыб отмечались изменения предпочитаемых участков в течение суток. Светлое время лещи проводили в более глубоком восточном расширении, в вечерние часы переходили в мелководное, с зарослями мягкой водной растительности западное расширение. Наиболее предпочитаемые рыбами в ночное время участки совпадали с местами концентрации моллюсков, которые доминировали в питании лещей. Слабое использование хирономид было связано с зарыванием их основной массы глубоко в детрит, что уменьшало их доступность. К концу июня регулярные суточные горизонтальные миграции прекратились, что было связано с сильным развитием в западном расширении зарослей, отрицательно сказавшимся на эффективности питания бентофагов. Известны регулярные суточные горизонтальные миграции леща в Рыбинском водохранилище. Перемещения по линии русло - мелководья на раннем этапе формирования этого водоема позволяли бентофагам использовать пищевые ресурсы обоих биотопов (Поддубный, 1971). В настоящее время улучшение условий питания па русловых участках привело к значительному сокращению диапазона суточных горизонтальных миграций леща (Линник, Герасимов, 1984), а в высококормном оз. Плещееве подобные регулярные перемещения вовсе не зарегистрированы (Малинин, Линник, 1983).
В прудах не было отмечено различий в предпочитаемых рыбами участках в течение суток. Как и в обвалованном водоеме, для лещей было характерно стабильное предпочитание мест с максимальными концентрациями основных объектов питания - личинок хирономид и ручейников в 1983 г., хирономид и моллюсков в 1984 г. В 1983 г. рыбы избегали потребления массовых групп моллюсков, предпочитая им значительно уступавших моллюскам по биомассе, но распределенных более агрегированно личинок ручейников.
Осваиваемые мечеными лещами участки водоемов сильно перекрывались (до 80%), но, как правило, расстояния между рыбами были не менее 2 м, и посещение одних и тех же точек перекрывавшихся участков разными особями происходило не синхронно, что свидетельствует об отсутствии стайности у исследуемых экземпляров в данной ситуации. На других видах рыб показано, что диапазон проявления стайного поведения в зависимости от условий среды может сильно изменяться (Helfman, 1984).
Распределение частот нахождения рыб с передатчиками в разных точках осваиваемой акватории подчинялось закону отрицательного биномиального распределения, что связано с экологической гетерогенностью водоемов. Степень неравномерности использования лещами акватории повышалась по мере привыкания рыб к водоемам (табл. 2).

Таблица 2. Показатели неравномерности посещения лещами разных участков прудов
* Отмечены статистически достоверные значения коэффициента Ллойда
Таким образом, после первоначального периода поисковых перемещений рыбы "знакомятся" с водоемом и в дальнейшем посещают только участки с субоптимальными сочетаниями факторов среды, т. е. характер освоения особями биотопа становится более упорядоченным. Скорее всего, и в естественных условиях процесс освоения рыбами новых "домашних" участков идет таким же путем.
Изменения тактики освоения пространства накладывают отпечаток на суточную динамику двигательной активности лещей. В начальный период поисковых перемещений средние скорости плавания довольно высоки (около 1,8 cм/с) и в течение суток изменяются незначительно. В дальнейшем на фоне общего снижения двигательной активности (скорость 0,5 cм/с) четко проявляются 2 - 3 пика, когда среднечасовые скорости плавания достигают 2,0 - 2,5 см/с. В обвалованном водоеме пики двигательной активности отсутствовали, но было отмечено повышение подвижности лещей в ночные часы (рис. 1). Существенные отличия ритмики двигательной активности рыб в различных водоемах могут быть связаны как с индивидуальной изменчивостью (Eriksson, 1978; Oishi et. al., 1984), так и с средовыми влияниями (Devine, Shiozava, 1984). Вариационным рядам скоростей плавания лещей во всех водоемах была свойственна сильная положительная асимметричность и перерассеянность (рис. 2), что свидетельствует о стремлении рыб к минимизации энергетических затрат на перемещения.
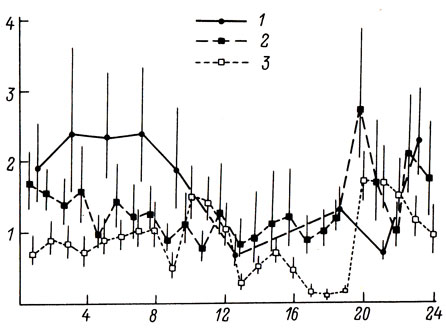
Рис. 1. Суточная ритмика двигательной активности лещей в обвалованном водоеме (1) и в пруду в 1983 г. (2) и 1984 г. (3). По оси ординат - средние геометрические скорости плавания и их доверительные интервалы (P=0,95), см/с; по оси абсцисс - время, часы
Из результатов проведенного исследования можно сделать следующие выводы.
1. Характер суточных горизонтальных перемещений половозрелого леща в мелководных искусственных водоемах зависит от степени развития мягкой водной растительности и разнообразия глубин водоема.
2. После выпуска леща в незнакомый водоем первые несколько суток рыбы перемещаются по большей части акватории, затем площадь, осваиваемая каждой особью, и ее двигательная активность уменьшаются.
3. Рыбы предпочитают находиться на участках, характеризующихся максимальным обилием доминирующих в питании кормовых организмов.
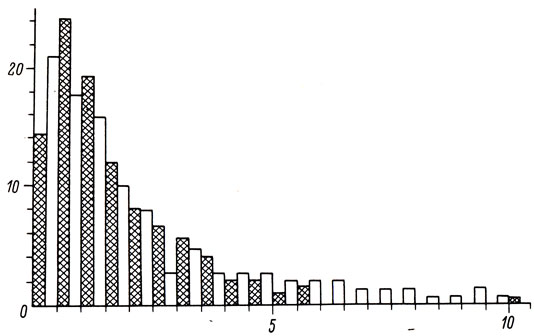
Рис. 2. Вариационные ряды скоростей плавания лещей в обвалованном водоеме (светлые столбцы) в пруду, 1983 г. (черные столбцы). По оси ординат - частота, %; по оси абсцисс - скорость, см/с
4. Небольшие среднесуточные скорости плавания (0,5 - 1,8 cм/с) и характер их варьирования свидетельствуют о стремлении рыб к минимизации затрат энергии на перемещения.
Антомонов Ю. Г. 1977. Моделирование биологических систем. Справочник. Киев.
Линник В. Д., Герасимов Ю. В. 1984. Поведение леща в нагульный период на высококормном участке Рыбинского водохранилища. - В сб.: Проблемы рыбохозяйственных исследований внутренних водоемов Северо-Запада европейской части СССР. Петрозаводск.
Малинин Л. К., Герасимов Ю. В., Линник В. Д., Саранчов С. И. 1983. Особенности нагульных перемещений леща в водоемах различного типа. - В сб.: Механизмы поведения. Материалы III Всесоюзн. конф. по поведению животных. М.
Малинин Л. К., Линник В. Д. 1983. Плотность и пространственное распределение массовых видов рыб в оз. Плещееве. - В сб.: Функционирование озерных экосистем. Рыбинск.
Пианка Э. 1981. Эволюционная экология. М.
Поддубный А. Г. 1971. Экологическая топография популяций рыб в водохранилищах. Л.
Поддубный А. Г., Малинин Л. К., Спектор Ю. И. 1979. Биотелеметрия в рыбохозяйственной науке и практике. М.
Саранчов С. И. 1984. Применение биотелеметрической техники в рыбохозяйственных исследованиях. - Обзорн. информ. ЦНИИТЭИРХ.
Шилов А. И. 1977. Эколого-физиологические основы популяционных отношений у животных. М.
Devine M. G., Shiozava D. K. 1984. Littoral heterogeneity and diel behavior of white bass (Morone chrysops) and carp (Cyprinus carpio) in Utah Lake, Utah. - Great Basin Naturalist, 44, 3.
Eriksson L. O. 1978. Nocturnalism versus diurnalism: dualism within fish individuals. - In: Thorpe J. E. (ed) Rhytmic activity of fishes. London.
Helfman G. S. 1979. Twilight activities of yellow perch, Perca flavescens. - J. Fish Res. Bd. Can., 36, 2.
Helfman G. S. 1984. School fidelity in fishes: the yellow perch pattern. - Animal Behav., 32, 3.
Mackay W. C., Craig J. F. 1983. A comparison of four systems for studying the activih of pike and perch. - Proc. 4th Int. Wildlife Biotelemetry Conf.
Oishi T., Shiigi C., Ueda-Kozai M., Tokura H. 1984. Circadian activity rhythm in a tropical fish, kissing gourami.- Abstr. 10 th Int. Congress of Biometeorology, Tokyo.
Stasko A. B., Pincock D. G. 1977. Review of underwater biotelemetry, with emphasis on ultrasonic techniques. - J. Fish Res. Bd.Can., 34, 9.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'