Морские биологические ресурсы
Биологическое обоснование промысловой меры аркто-норвежсной трески и пикши (Канд. биол. наук Низовцев Г.П., канд. биол. наук Ковцова М.В. и Третьяк В.Л., ПИНРО) (УДК 597.562-152.6(268.43/45))
Характерная особенность современного рыболовства в Баренцевом море - общность запасов основных промысловых рыб для советской и норвежской экономических зон - далее СЭЗ и НЭЗ. Как известно, в Баренцевом море, особенно в СЭЗ, обитают в основном неполовозрелые треска и пикша. По мере созревания треска и пикша уходят на нерестилища в зону Норвегии, которая отчасти является и местом зимовки половозрелых и неполовозрелых рыб. После нереста треска мигрирует в Баренцево море на нагул и создает там динамичные скопления, которые советским флотом почти не облавливаются. Отнерестившаяся пикша возвращается в СЭЗ в незначительном количестве (в среднем около 10%).
Указанные особенности распределения запасов трески и пикши вызывают определенные затруднения в рациональном их использовании, поскольку различная возрастная структура уловов диктует Норвегии и СССР прямо противоположную политику при выработке мер регулирования рыболовства посредством размера ячеи тралов и промысловой меры на треску и пикшу. Норвегия заинтересована в том, чтобы сохранить неполовозрелую молодую рыбу, дать ей возможность вырасти до крупных размеров, когда она достигнет половой зрелости и станет доступной для норвежского промысла. Поэтому уже в течение ряда лет норвежские ученые настаивают на увеличении размера ячеи тралов до 135-155 мм. Однако увеличение ячеи как мера регулирования промысла имеет существенные недостатки.
Цель настоящей работы - оценка влияния различных мер регулирования на состояние эксплуатируемых популяций трески и пикши в Баренцевом море, результативность их и попытка биологического обоснования оптимальной промысловой меры на треску и пикшу как одной из мер регулирования промысла.
В настоящее время регулирование промысла осуществляется по трем основным направлениям: размеру ячеи орудий лова и промысловой мере на добываемую рыбу, квотированию величины ежегодного вылова по видам рыб, комплексу мер по охране молоди рыб и нерестующей рыбы.
В Баренцевом море началом регулирования промысла с помощью определенной ячеи тралов следует считать 1946 г., когда была выработана конвенция по рыболовству в Северо-Восточной Атлантике. Конвенция была ратифицирована в 1959 г. С этого года на промыслах донных рыб в Баренцевом море была установлена ячея тралов 110 мм. Затем в 1967 г. ячея была увеличена до 120 мм, установлена промысловая мера на треску и пикшу соответственно . 34 и 31 см, в 1981 г. ячею увеличили до 125 мм, а промысловую меру для трески и пикши до 39 и 35 см. В 1982 г. промысловая мера на треску и пикшу без какого-либо биологического или экономического обоснования была увеличена по решению Смешанной советско-норвежской комиссии по рыболовству до 42 и 39 см. В 1983 г. Норвегия в одностороннем порядке ввела в своей зоне ячею 135 мм.
Для того чтобы оценить влияние постепенного увеличения ячеи и промысловой меры на популяции трески и пикши, рассмотрим многолетнюю динамику размерно-возрастной структуры популяций, численности ежегодного пополнения, рыболовного эффекта (вылова) в связи с динамикой коэффициента промысловой смертности F.
Несмотря на поэтапное увеличение ячеи траловых мешков и промысловой меры на добываемую рыбу, процесс нарушения возрастной структуры популяции продолжается. Особенно это касается правой части возрастного ряда, т. е. нерестового запаса популяций.
Исследования показали, что на фоне истощения производителей трески и пикши при высокой интенсивности промысла дефицит пополнения промысловой части стада не уменьшается, а дефицит рыболовного эффекта особенно за последнее десятилетие заметно увеличился (рис. 1 и 2).
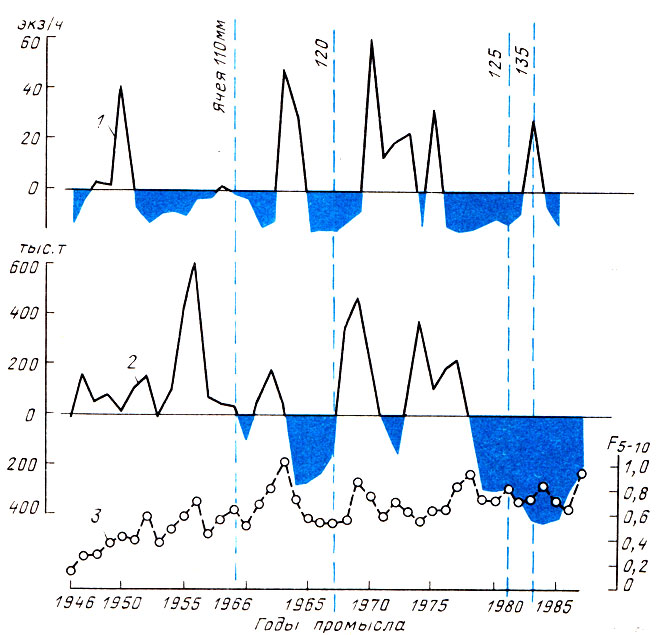
Рис. 1. Динамика некоторых показателей эксплуатации трески аркто-норвежской популяции: 1 - численность ежегодного пополнения (средний улов, в экз. на час траления, молоди в возрасте 2+); 2 - вылов всеми странами в отклонениях от средней многолетней (719 тыс. т); 3 - коэффициент промысловой смертности (5-10)
Для суждения о том, к чему приведет дальнейшее увеличение размера ячеи и промысловой меры, следует обратиться к опыту регулирования промысла трески и пикши в исландских водах. Как известно, там одновременно с установлением 200-мильной рыболовной зоны в 1976 г. была введена ячея 155 мм, промысловая мера 55 см. Применяя до настоящего времени указанную ячею, исландцы тем самым провели своего рода долговременный натурный эксперимент влияния крупной ячеи на структуру популяции и результативность промысла. Сравнительные данные о возрастном составе популяции трески исландского стада за ряд лет (Jonsson, 1955, 1979, 1982; Schopka, 1984; Hafrannsoknastofnun fjolrit, 1988) свидетельствуют о том, что применение на промысле ячеи 155 мм не оказало положительного влияния на возрастную структуру популяции. За годы, прошедшие с начала применения ячеи 155 мм, увеличился дефицит ежегодного пополнения промысловой части стада молодью. Рыболовный эффект также уменьшился. Аналогичная ситуация наблюдается с пикшей исландского стада.
Из приведенных выше материалов следует, что увеличение размера ячеи и промысловой меры на добываемую треску и пикшу без снижения интенсивности рыболовства не дает положительных результатов. Например, опыт 12-летнего промысла трески в районе Исландии ячеей 155 мм показал, что возрастная структура нерестовой части популяции не только не восстановилась, но продолжает истощаться. Если в 1977 г. рыбы в возрасте более 8 лет составляли в популяции 13,6%, то в 1988 г. уже только 2,3% (Anon., 1988). Аналогичный процесс идет в популяциях баренцевоморской трески и пикши: в 1946-1950 гг. в советских уловах было зарегистрировано трески в возрасте более 8 лет 8,4 %, в 1981 - 1986 гг.- всего 1,0%. Пикши старше 7 лет в 1946-1950 гг. было в уловах 30,0%, в 1981 - 1987 гг.-4,1 %. Происходит это потому, что увеличенная ячея селективно отбирает из популяции крупных рыб, являющихся наиболее продуктивными производителями, имеющими значительно большую индивидуальную плодовитость, чем впервые нерестующие рыбы. В результате популяционная плодовитость начинает прогрессирующе снижаться.
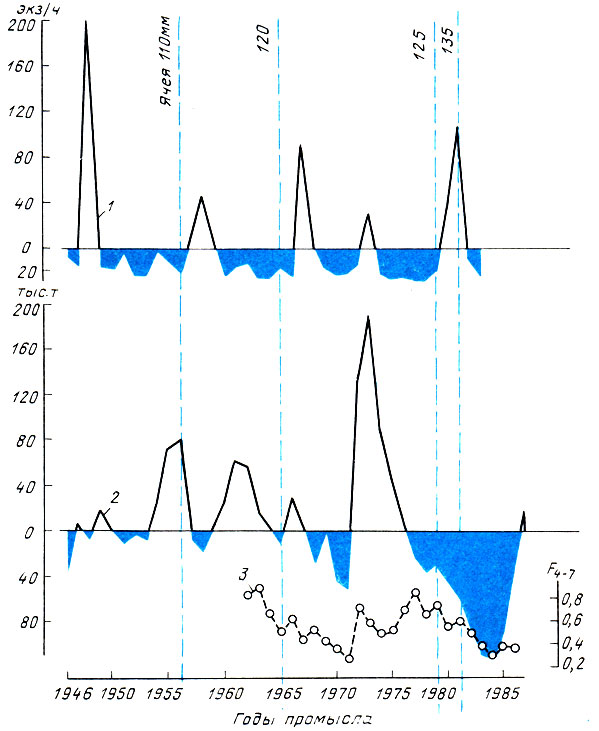
Рис. 2. Динамика некоторых показателей эксплуатации баренцевоморской пикши: 1 - численность ежегодного пополнения (средний улов молоди в возрасте 2+) в отклонениях от средней многолетней (131 тыс. т); 2 - вылов всеми странами в отклонениях от средней многолетней (131 тыс. т); 3 - коэффициент промысловой смертности (4-7)
По последним исследованиям, индивидуальная абсолютная плодовитость трески колеблется от 170 тыс. до 18 млн икринок. В 1987 г. самые высокие значения индивидуальной абсолютной плодовитости наблюдались у рыб в возрасте 9-13 лет (Низовцев, Пахомова, Ярагина, 1988). Однако нерестовое стадо баренцевоморской трески непрерывно омолаживается, поскольку убыль рыб старших возрастных групп из-за чрезмерной интенсивности промысла не компенсируется более молодыми производителями, что в конечном итоге приводит к минимальным значениям популяционной плодовитости и к увеличению частоты появления малочисленных поколений. Так, с 1945 по 1967 г. (за 23 года), когда нерестовое стадо трески находилось в относительно благополучном состоянии, таких поколений появилось 5, а с 1968 по 1988 г. (за 20 лет) их было уже 12.
Популяция баренцевоморской пикши с 1948 по 1967 г. (за 20 лет) дала 6 малоурожайных поколений, а с 1968 по 1988 г. (за 21 год) - 12. Это следствие того, что в нерестовом стаде стала преобладать пикша младших возрастных групп, индивидуальная плодовитость которой значительно ниже, чем у рыб старших возрастов. Популяционная плодовитость пикши в последние годы (1978-1988) находилась на минимально допустимом уровне (Kovtsova, 1989).
Отрицательное воздействие увеличенной ячеи активных отцеживающих орудий лова сказывается не только на нерестовой, но и на неполовозрелой части популяции. Проходящая через ячею рыба подвергается интенсивному физическому и физиологическому травмированию, в результате которого гибнет.
О травматической гибели рыб после прохождения через ячею трала свидетельствуют как наблюдения ПИНРО, проведенные в 1977-1988 гг., так и многие предшествующие исследования, описанные в литературе. Например, наблюдения из гидростата за движением трала по дну показали, что в кутке движущегося трала ячея имеет форму сильно вытянутого ромба и через такую ячею рыбе пройти весьма трудно (Логунов, 1955). Следовательно, делает вывод автор, рыба выходит из трала при поднятии на поверхность и не может быть жизнеспособной, поскольку длительное время подвергалась сдавливанию в мешке как при движении трала по дну, так и при поднятии с глубины на поверхность.
Подобные результаты были получены Г. Оттерлиндом (Otterlind, 1960) в отношении молоди балтийской трески. По его наблюдениям, выживаемость трески длиной 31-35 см составила 50 %, длиной 10-15 см - всего 5 %. Причиной гибели молоди трески Г. Оттерлинд считает смену внешнего давления при поднятии рыбы из придонных слоев на поверхность моря.
Данные, полученные при изучении выживаемости рыб в экспериментальных условиях, показали, что те особи, которые уходят через ячею трала, погибают из-за чрезмерного накопления в крови молочной кислоты (Templemann, 1963; Bagge, 1970).
Подводные наблюдения из ПА "Тетис" и фотосъемка (Заферман, Серебров, 1989) подтверждают тот факт, что во время траления выход рыб через ячею возможен только в узкой зоне ее максимального раскрытия на участке сетного полотна перед накопленной массой улова. Внутри мешка турбулентные потоки лишают рыбу ориентации, прижимают к дели, заставляют выходить через ячею в разных положениях, что приводит к ее травмированию. Выход рыбы через ячею хвостом вперед неизбежно влечет за собой травмирование плавников, жаберных крышек и чешуи. Однако и при активном выходе головой вперед рыбы также могут быть сильно травмированы обтекающим трал потоком воды, который прижимает рыбу к дели в момент выхода При этом могут быть повреждены даже мелкие особи, которые при отсутствии потока могли бы свободно пройти через ячею.
Исследования показали, что после прохождения трала на грунте имеются погибшие особи пикши. Их число и размерный состав примерно соответствовали численности и размерам рыб, отсеиваемых из мешка через ячею и собираемых уловителем.
Количественными оценками гибели трески, прошедшей через ячею тралов, сотрудники ПИНРО пока не располагав ют. Однако среди живых рыб, отмеченных в протравленной полосе, часто можно было видеть особей трески со следами пребывания в трале: содранная чешуя по периметру обхвата тела, глубокие отпечатки сетей на боковых поверхностях тела. Снижение количества маломерных рыб в трале при увеличении размера ячеи создает лишь иллюзию сохранности молоди. При увеличении ячеи увеличивается количество рыбы, проходящей через нее; многие рыбы могут проходить через ячею неоднократно. Соответственно возрастает и их травматизм. При этом многие рыбы погибают непосредственно от физических и физиологических травм, или сильно ослабленные они становятся легкой добычей для хищников. Косвенным доказательством этого является тот факт, что увеличение ячеи на промысле тресковых не дало ожидаемого положительного эффекта ни в Баренцевом море, ни в районе Исландии и, более того, повлекло за собой дальнейшее разрушение естественной структуры популяций трески и пикши.
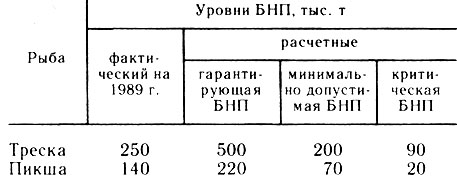
Таблица 1
Имеется более эффективная мера регулирования, основанная на оптимизации интенсивности промысла, в частности на оптимизации биомассы половозрелых рыб. Расчеты уровней биомассы нерестовой популяции (БНП) в Баренцевом море, выполненные на основе данных популяционной плодовитости по методу, предложенному В. П. Серебряковым (Serebryakov, 1988), дали следующие величины (табл. 1).
Для организации рационального промысла необходимо увеличить биомассу нерестовых популяций до гарантирующей величины и затем поддерживать на достигнутом уровне. Это осуществляется путем изменения интенсивности рыболовства F, которая рассчитывается ежегодно. По отношению к эксплуатируемой популяции, имеющей тенденцию к депрессии, эта мера на практике означает различную степень снижения интенсивности промысла (уменьшение ОДУ). Для популяции, находящейся в депрессии, уменьшение вылова должно быть значительным, вплоть до полного запрета лова на ряд лет, необходимых для восстановления БНП и общей численности популяции.
В результате снижения интенсивности рыболовства начинается постепенный рост биомассы половозрелых рыб, которая уже через несколько лет достигает оптимальной величины и далее изменяется незначительно. Увеличение биомассы нерестовой популяции до гарантированного уровня приводит к увеличению воспроизводительной способности, т. е. популяционная плодовитость поднимается до уровня, при котором обеспечивается появление урожайных поколений при благоприятных условиях выживания. Это исключает появление малочисленных поколений из-за низкой численности половозрелой части популяции и обеспечивает на перспективу стабильно высокий рыболовный эффект с затратой значительно меньших рыболовных усилий, т. е. приводит промысел к состоянию рационального.
Приняв главной мерой регулирования промысла предлагаемую, т. е. общий допустимый улов (ОДУ) и национальные квоты, ежегодная величина которых рассчитана с учетом создания и постоянного сохранения гарантирующих нерестовых запасов (такой численности половозрелых рыб, которая гарантирует популяцию от появления неурожайных поколений из- за недостатка производителей), второй важнейшей мерой системы регулирования должна стать охрана молоди рыбы от преждевременного вылова. В противном случае может быть сведен на нет эффект от первой, главной, меры.
Реализация последней рыбоохранной меры требует четкого обоснования понятия "молодь рыбы". В понимании, что такое молодь того или иного вида рыбы, у исследователей до настоящего времени нет единой точки зрения. Теоретическая основа здесь одна - общее положение К. М. Бэра (1960) и Н. Я. Данилевского (1875) о поддержании воспроизводительной способности популяции на достаточно высоком уровне и в связи с этим - представление о целесообразности охраны от вылова рыб, не достигших половой зрелости. Согласно этому представлению, наименьшую промысловую меру на рыб с длительным жизненным циклом принимают равной длине, при которой рыба становится половозрелой, или средней длине половозрелых особей. Можно говорить, что такой принцип установления минимального размера разрешенной к вылову рыбы в первом приближении удовлетворяет широко распространенному в ихтиологической литературе мнению о необходимости предоставления возможности каждой рыбе, по крайней мере, один раз участвовать в нересте. Однако это мнение никак не обосновано количественно и применимо далеко не к каждому виду рыб. Это положение справедливо, по-видимому, прежде всего по отношению к таким видам рыб, у которых период полового созревания наступает в сравнительно раннем возрасте и при небольших длине и массе тела преимущественно в одном возрасте. Вряд ли оно пригодно при определении минимальной промысловой меры для видов с длительным жизненным циклом, к каким относятся треска и пикша, имеющим высокий темп роста, значительные индивидуальные колебания длины и массы тела, у которых особи задолго до наступления половой зрелости достигают значительной длины и массы и представляют собой отличную товарную рыбу. Определенный объем вылова такой неполовозрелой рыбы, рассчитанный с учетом сохранения гарантированного нерестового запаса, вполне возможен и оправдан биологически и экономически. При обосновании минимальной промысловой длины нужно исходить из принципа сохранения естественной структуры популяции, т. е. осуществлять промысел таким образом, чтобы промысловое изъятие было пропорционально численности возрастных классов, составляющих данную популяцию, за исключением молоди. Каких же рыб (до какого размера) следует причислять к молоди?
По нашему мнению, к молоди нужно относить особей до наступления той длины, с которой начинается половое созревание в популяции. Например, в связи с неравномерным ростом особей одного поколения, а также с различным ростом рыб разных поколений половое созревание у пикши очень растянуто (Ковцова, 1987). В одни годы доля половозрелых рыб длиной 36-40 см составляет 25- 27%, длиной 41-45 см - более 40% (1982, 1983 гг.), в другие годы доля половозрелых особей длиной 36-40 см - менее 1 % (1978 г.). Особенно сильно ускоряется темп роста особей бедных поколений, когда численность промысловой части популяции низкая (табл. 2).
По многолетним данным, наступление половой зрелости у пикши начинается в возрасте 3 лет при длине 35 см и заканчивается в возрасте 12-13 лет при длине 80 см (Ковцова, 1987). В массовом количестве созревают рыбы в возрасте 5-7 лет при длине 51-60 см.
Таким образом, раннее массовое созревание пикши в возрасте 3-4 лет наблюдается довольно часто, поскольку бедные поколения появляются значительно чаще, чем богатые. Следовательно, правомерно считать молодью пикши всех особей того размера, при котором начинается половое созревание в популяции, т. е. при длине 35 см включительно. Все экземпляры, достигшие длины 36 см и более, следует относить к особям промысловой длины.
Треска созревает медленнее, чем пикша, однако первые половозрелые особи также появляются среди 3-годовиков при длине 36-40 см. Массовое созревание происходит в возрасте 7-9 лет при длине 70-80 см и заканчивается в возрасте 12-13 лет (см. табл. 2). Поскольку количество половозрелой трески в размерной группе 36-40 см значительно меньше по сравнению с пикшей (по средним многолетним данным оно составляет 0,3% против 4,6% у пикши), целесообразно всю треску указанных размеров (3(6-40 см) относить к молоди и минимальной промысловой длиной трески считать 41 см. С 1989 г. наблюдается тенденция снижения численности промыслового стада трески и пикши в связи с очень слабым пополнением его бедными поколениями 1985-1987 гг., следовательно,в 1990 г. половое созревание особей должно ускориться, что приведет к значительному увеличению доли половозрелых рыб в возрасте 3- 4 лет.
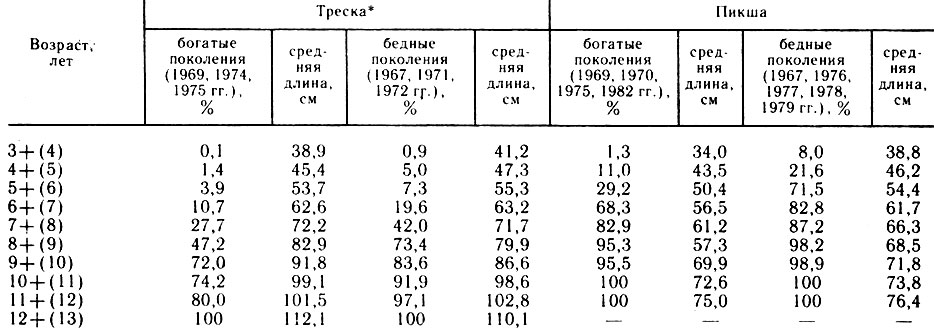
Таблица 2
*(По В. П. Пономаренко, И. Я. Пономаренко, Н. А. Ярагиной, 1985.)
Исследованиями ПИНРО установлено, что в годы высокой численности пополнения допустимый прилов молоди трески и пикши, который не нанесет ущерба воспроизводительной способности популяций, составляет 15 %. По-видимому, в годы слабого пополнения и низкой численности промыслового стада величина прилова маломерных рыб будет более низкой. Следовательно, допустимая величина прилова молоди должна быть величиной переменной и зависеть от численности пополнения. Отсюда следует, что расчет допустимых приловов маломерной трески и пикши должен производиться исходя из величины пополнения и квот вылова.
Таким образом, если величина допустимого прилова молоди будет рассчитана правильно и отдельно для разных по численности пополнения лет, то появится больше шансов для того, чтобы значительная часть быстрорастущих особей отнерестилась не 1 раз в жизни, а 2 и 3 раза.
До настоящего времени промысловая мера на треску и пикшу соответствовала точке 50 %-ного отсева на кривой селективности для регламентированного размера ячеи. Такой подход, когда промысловая мера на рыбу является производной от размера ячеи, противоречит здравому смыслу и необоснован ни биологически, ни экономически. Так, Норвегия с 1 января 1990 г. в своей экономической зоне увеличивает минимальную промысловую длину на треску и пикшу до 47 и 44 см соответственно, а два года подряд (1987-1988) настаивала на увеличении размера ячеи по всему Баренцевому морю до 155 мм. Однако единственной причиной для такого увеличения является снижение интенсивности промысла в южной части Баренцева моря (в СЭЗ). Логически же необходимо, исходя из биологически обоснованной промысловой длины на треску и пикшу (41 и 36 см), подобрать такую ячею трала, которая удерживала бы рыбу промысловой длины максимально, а молодь - в пределах рассчитанной допустимой величины.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'