Эмбриональное развитие сига-пыжьяна coregonus lavaretus pidschian (Gmel.) в условиях ЦЭС ГосНИОРХ "Ропша" (Г. А. Головкова)
Одним из направлений повышения рыбопродуктивности водоемов и более эффективного использования их кормовой базы является метод выращивания поликультуры рыб, различающихся по спектрам питания. В условиях Северо-Запада целесообразно вводить в поликультуру сиговых рыб (Руденко, 1975, 1978). В качестве планктофага успешно используется пелядь, в качестве бентофага перспективен сибирский сиг-пыжьян. Последний рано переходит на исключительно бентосное питание, имеет хороший темп роста, быстро созревает. На начальном этапе работ по рыбоводному освоению пыжьяна проводилось изучение его эмбрионального развития в новых условиях.
Материал и методика. Материалом для работы служила икра пыжьяна, полученная от производителей, созревших на ЦЭС ГосНИОРХ "Ропша". Инкубация икры проходила на ЦЭС "Ропша" в литровых аппаратах Вейса. Расход воды колебался от 1,5 до 2,2 л/мин, содержание кислорода составляло 7,2 - 14,5 мг/л, углекислого газа - 0,8 - 3,2 мг/л, pH - 7,8 - 8,8. Среднемесячная температура воды колебалась от 2,2 до 5,2°С.
Наблюдения за развитием зародышей проводились на икре, фиксированной 4%-ным формалином, и на живых эмбрионах. В первые полтора часа после оплодотворения фиксации производились через 1, 2, 5 и 10 мин. Далее до возраста зародышей в одни сутки интервал между фиксациями постепенно увеличивали до 1 часа. В возрасте 1 - 3 суток интервал составлял 6 час., а в дальнейшем был увеличен до 1 - 2 суток. Всего исследовано 91 фиксированных проб икры пыжьяна. В основу подразделения эмбрионального развития на этапы принята периодизация, предложенная С. Г. Крыжановским с соавторами (1953) и И. И. Смольяновым (1957).
Результаты исследований. Эмбриональный период развития пыжьяна был подразделен на 8 этапов. Описание эмбриогенеза приводится по этапам и стадиям.
Средний диаметр неоплодотворенной икринки пыжьяна составляет 2,08±0,113 мм. Икра желто-оранжевого цвета, мелкие жировые капли диаметром около 0,15 мм концентрируются на анимальном полушарии, остальную часть яйца занимает однородный по консистенции желток (рис. 1 а). Вследствие такого расположения жировых капель, имеющих меньший удельный вес, яйцо ориентировано анимальным полюсом вверх, что способствует улучшению газообмена зародыша (Соин, 1968).
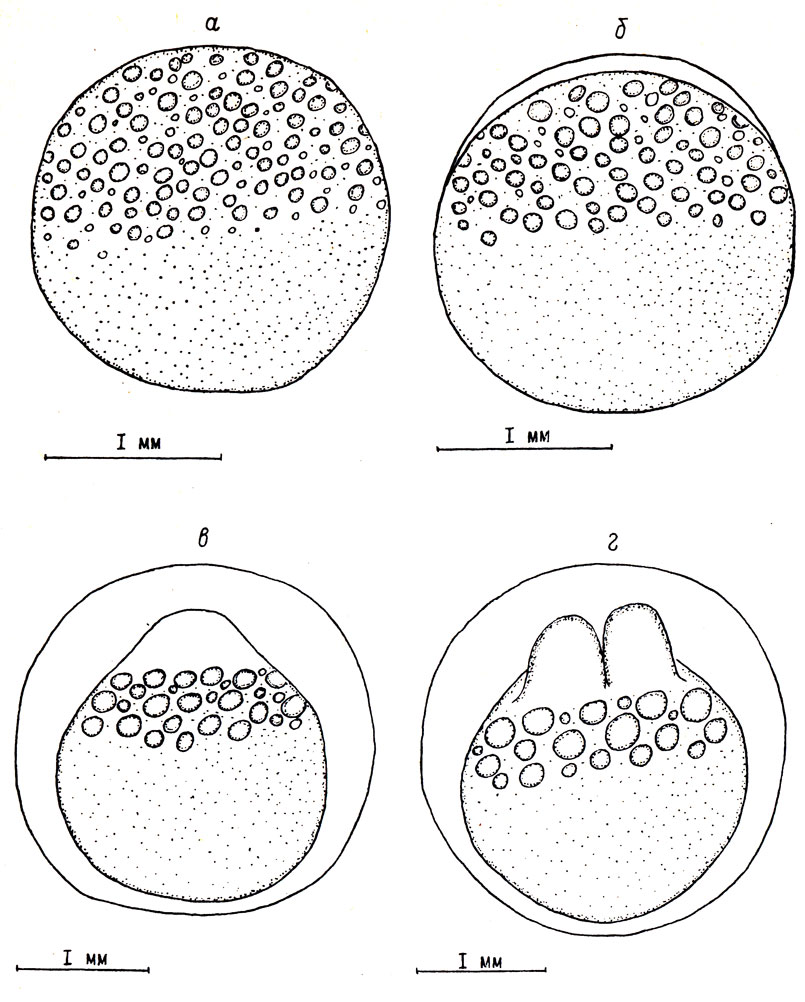
Рис. 1. Начальные стадии эмбрионального развития пыжьяна: а - неоплодотворенная икринка; б - возраст 10 мин, образование перивителлинового пространства; в - возраст 14 час, сформированный бластодиск; г - возраст 16 час, стадия двух бластомеров
На первом этапе развития образуется перивителлиновое пространство, за счет чего увеличивается объем икринки. Одновременно происходит образование бластодиска. Этап начинается с момента оплодотворения и заканчивается с началом дробления. При средней температуре воды 2,4° этап продолжается 16 час.
Через 1 мин после оплодотворения видимых изменений в строении яйца не происходит. Спустя 5 мин на анимальном полюсе начинает образовываться перивителлиновое пространство, которое в дальнейшем увеличивается в объеме. Через 10 мин после осеменения диаметр икринки достигает 2,16 мм, перивителлиновое пространство занимает 10,4% ее объема (см. рис. 1, б).
Процесс набухания заканчивается через 6 час. К этому времени диаметр икры увеличивается на 32,2% и достигает 2,75 мм. Объем икры возрастает в 2,3 раза.
Одновременно происходит образование бластодиска. Через 5 час. после оплодотворения на анимальном полюсе яйца появляется холмик цитоплазмы. К концу этапа бластодиск сформирован: его высота составляет 0,5 мм, диаметр основания - 1,05 мм. Жировые капли располагаются в несколько рядов под основанием бластодиска (см. рис. 1, в).
На втором этапе развития происходит дробление бластодиска. Этап начинается с появления первой борозды дробления и заканчивается образованием эпителиальной бластулы, клетки которой достигли минимальных размеров. При средней температуре воды 2,8° этап длится с 16 час после оплодотворения до возраста зародыша 8 суток.
Первая борозда дробления появляется у некоторых икринок спустя 16 час после оплодотворения. Средние размеры бластомера составляют 1,10×0,60×0,45 мм (см. рис. 1, г). Вторая борозда, расположенная перпендикулярно первой, образуется через 20 час после оплодотворения. К этому времени каждый из двух бластомеров делится пополам. Средние размеры бластомера уменьшаются до 0,60×0,50×0,45 мм. Жировые капли, подстилающие бластомеры, сливаются в более крупные, диаметром 0,18 мм (рис. 2, а).
Через 30 час все икринки находятся на стадии четырех бластомеров. Спустя 42 час с момента оплодотворения образуется 8 бластомеров, расположенные в один слой. Средние размеры бластомера составляют 0,40×0,25×0,45 мм.
Стадия 16 бластомеров, расположенных в два слоя, наблюдается в возрасте 2 суток (сумма тепла 4,8°). Средний диаметр бластомера равен 0,175 мм. В возрасте 60 час происходит очередное деление и образуются 32 бластомера.
При последующих делениях бластомеров происходит увеличение количества и уменьшение размеров клеток. Этап дробления заканчивается стадией эпителлиальной бластулы в возрасте 8 суток (сумма тепла 22,4°). Диаметр зародыша у основания составляет 1,05 мм, высота его - 0,45 мм. Средний диаметр бластомера снижается до 0,014 мм и в дальнейшем не уменьшается (см. рис. 2, б).
На третьем этапе происходит гаструляция. Этап охватывает период с начала обрастания желтка бластодермой до образования желточной пробки. При средней температуре воды 2,6° этап длится с 9-х по 14-е сутки.
На 9-е сутки развития (сумма тепла 24,6°) начинается обрастание желточного мешка бластодермой. Средний диаметр бластодиска увеличивается до 1,55 мм. В наиболее утолщенной части бластодиска образуется "краевой узелок".
В возрасте 11 суток (сумма тепла 29,1°) бластодермой обрастает 1/2 желтка. На границе с желтком видна светлая полоска перибласта, подстилающего бластодерму. "Краевой узелок" удлиняется и превращается в "краевой язычок", где образуются зародышевые пласты экто-, мезо- и энтодермы. В дальнейшем здесь будут формироваться головной, туловищный и хвостовой отделы зародыша.
На 13 - 14-е сутки развития (сумма тепла 37,6°) наблюдается стадия "желточной пробки".
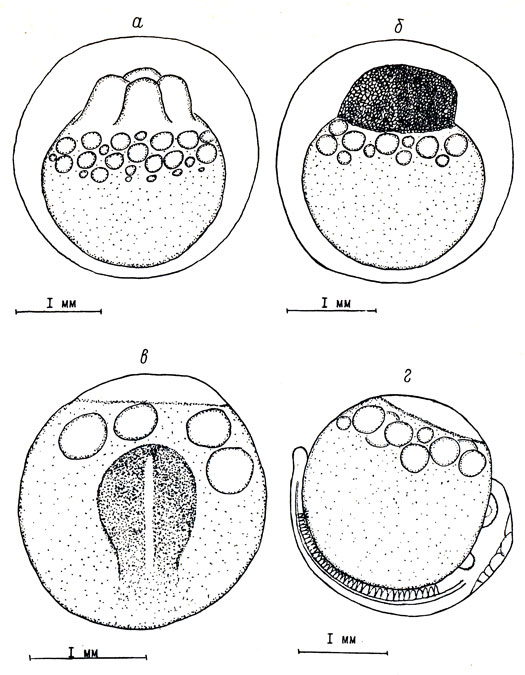
Рис. 2. Второй, третий, четвертый этапы эмбрионального развития пыжьяна: а - возраст 1 сутки, стадия четырех бластомеров; б - возраст 8 суток, стадия эпителиальной бластулы; в - возраст 15 суток, начало формирования головного и туловищного отделов. Оболочка снята; г - возраст 26 суток, образование хвостовой почки. Оболочка снята
Четвертый этап развития - закладка головного и туловищного зачатков. В это время происходит формирование основных органов зародыша: хорды, нервной трубки, мускулатуры. Появляется зачаток головного мозга. В головном отделе зародыша формируются глазные пузыри, слуховые плакоды. Начинается сегментация туловищной мускулатуры. Этап начинается с обособления головного отдела зародыша и заканчивается образованием хвостовой почки. При средней температуре воды 2,5° он продолжается с 15-х по 26-е сутки.
В возрасте 15 суток (сумма тепла 40,9°) на стадии "желточной пробки" у зародыша формируются расширенный головной и более узкий туловищный отделы. В туловищной части образуются нервная борозда и хорда. Длина зародыша достигает 1,0 мм. Продолжается слияние и укрупнение жировых капель, их средний диаметр равен 0,35 мм (см. рис. 2, в).
На 17-е сутки развития (сумма тепла 46,9°) при замыкании бластопора у зародышей формируется хвостовой отдел. В головном отделе начинается закладка глазных пузырей. В теле зародыша образуется 3 пары туловищных сомитов.
На 18 - 19-е сутки (сумма тепла 51,2°) общая длина зародыша составляет 2,8 мм, ширина головного отдела - 0,8 мм. Зародыш начинает выступать над желточной массой. В это время происходит формирование глазных пузырей и закладка слуховых плакод. Перед головным концом зародыша имеется перибластический синус, выполняющий гидростатическую функцию. У зародышей продолжается закладка туловищных сомитов, их число достигает 6 пар.
В возрасте 22 суток (сумма тепла 64,4°) длина зародыша составляет 3,0 мм. Хвостовой отдел становится более компактным. Число сомитов достигло 20 пар. Размеры перибластического синуса равны 1,41×0,28 мм. Глазные пузыри путем впячивания передней стенки преобразуются в глазные бокалы, в них начинается образование хрусталика. В головном мозгу произошла дифференцировка первого мозгового пузыря на передний, средний и промежуточный мозг.
В возрасте 26 суток (сумма тепла 71,4°) длина зародыша равна 3,3 мм. Число сомитов достигло 38 пар. В головном мозгу произошло увеличение среднего мозга. Началась сегментация второго мозгового пузыря - в нем образовалось 5 нейромеров. Слуховые плакоды преобразовались в слуховые капсулы. Длина глазных бокалов равна 0,37 мм, межглазничное расстояние - 0,35 мм, расстояние между глазными бокалами и слуховыми капсулами - 0,3 мм. Хвостовой конец приобрел каплевидную форму (см. рис. 2, г).
На пятом этапе происходит отчленение туловищно-хвостового и головного отделов от поверхности желтка. Этап начинается с отделения хвостовой почки от желтка и заканчивается с появлением на теле зародыша меланофоров. При средней температуре воды 2,7° длится с 27-го по 41-й день.
На 27-е сутки развития (сумма тепла 73,6°) хвостовая почка частично отделилась от поверхности желтка.
На 30-е сутки (сумма тепла 81,4°) длина зародыша равна 4,0 мм, длина отчлененного туловищно-хвостового отдела - 1,0 мм. Время от времени зародыш совершает им колебательные движения. В связи с этим роль перибластического синуса как гидростатического органа уменьшается, и он начинает редуцироваться. Размеры его составляют 0,88×0,13 мм. Впереди глаз появились обонятельные плакоды. Жировые капли слились в две крупные, диаметром 0,38 мм каждая. Число туловищных сегментов достигло 40 пар.
В возрасте 33 суток (сумма тепла 89,7°) длина зародыша составляет 4,3 мм, ширина головы - 0,9 мм. Обонятельные плакоды преобразовались в обонятельные ямки. Перибластический синус продолжает уменьшаться, его размеры составляют 0,75×0,12 мм. В желточном мешке имеется одна крупная жировая капля диаметром 0,79 мм и несколько мелких.
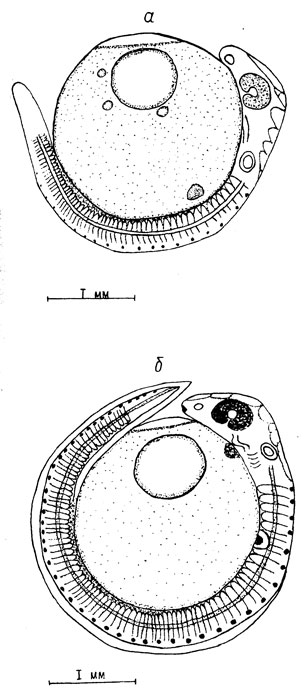
Рис. 3. Пятый и шестой этапы эмбрионального развития пыжьяна: а - возраст 41 сутки, начало пегментации; оболочка снята; б - возраст 54 суток, формирование ротовой воронки, кишки, жаберных дуг. Оболочка снята
В возрасте 41 суток (сумма тепла 113,9°) длина зародыша составляет 4,8 мм. Задняя часть туловищно-хвостового отдела длиной 1,7 мм отчленена от желтка. Начинается отчленение головного отдела зародыша. Происходит увеличение размеров глазных бокалов и слуховых капсул, размеры которых составляют соответственно 0,51×0,37 мм и 0,25×0,12 мм. Расстояние между ними равно 0,37 мм. Глаза зародышей слабо пигментированы. На желточном мешке по бокам тела зародыша на уровне 5-го сегмента образовались зачатки грудных плавников. Впереди на нижней стороне тела зародыша расположена сердечная трубка. На туловище в желточном мешке появились точечные меланофоры. Число туловищных сегментов достигло 56 пар (рис. 3, а).
Шестой этап характеризуется формированием системы сосудов и началом кровообращения. Происходит образование ротовой воронки, кишки, грудных плавников. Жировые капли сливаются в одну. При средней температуре воды 2,4°С этап длится с 42-х по 61-е сутки.
В возрасте 49 суток (сумма тепла 129,5°) длина зародыша составляет 5,4 мм. Число сегментов достигло 61 - 62 пар и в дальнейшем не увеличивается. Зародыш совершает колебательные движения отчлененной частью тела более активно. Пигментация тела стала интенсивнее: на спинной части туловища появились два ряда меланофоров. Тело зародыша обрамляет узкая плавниковая кайма.
На нижней поверхности головы формируется ромбовидный зачаток ротовой воронки, имеющей размеры 0,30×0,15 мм. Размеры глазных бокалов и слуховых капсул продолжают увеличиваться (соответственно 0,55×0,37 мм и 0,31×0,22 мм). Расстояние между ними составляет 0,37 мм.
В туловищной части зародыша появился зачаток кишки в виде сплошного тяжа клеток. Сердечная трубка приобрела изогнутую форму, справа от нее виден сосудистый мешочек. Грудные плавники переместились на тело зародыша, их основания расположены еще горизонтально.
У большинства зародышей все жировые капли слились в одну диаметром 1,25 мм.
В возрасте 54 суток (сумма тепла 143,6°) длина зародыша достигла длины окружности яйца - 6,53 мм. Зародыш совершает активные движения отчлененной частью тела. Голова полностью отделена от желточного мешка. Слуховая капсула переместилась вверх и приблизилась к заднему краю глаза. Расстояние между ними сократилось до 0,25 мм. Под слуховой капсулой сформировались жаберные дуги. Грудные плавники приобрели округлую форму, их центральная часть утолщена. Длина грудных плавников составляет 0,25 мм (см. рис. 3, б).
У всех зародышей жировые капли слились в одну, диаметр которой равен 1,25 мм. Длина желточного мешка 2,08 мм.
В возрасте 61 суток (сумма тепла 159,4°) длина зародыша достигает 7,2 мм. Усилилась пигментация тела: появились звездчатые меланофоры на верхней части головы и желточном мешке.
Размеры глазных бокалов и слуховых капсул продолжают увеличиваться и достигают соответственно 0,70×0,40 мм и 0,45×0,30 мм. Расстояние между ними уменьшается до 0,20 мм. В слуховой капсуле видны полукружные каналы и отолиты. Формирующиеся жаберные крышки еще не доходят до жаберных дуг. Грудные плавники приобрели подвижность, их длина 0,40 мм, основания расположены вертикально. В плавниковой кайме образуется хвостовая лопасть. Длина кишки равна 2,35 мм. Слева от кишки на желточном мешке располагается зачаток печени. Желточный мешок уменьшается, его размеры составляют 2,06×1,80 мм. На желточном мешке перед жировой каплей сохраняется остаток перибластического синуса.
Сердце состоит из трех отделов: венозного синуса, предсердия и желудочка. Из сердца кровь по жаберным дугам поступает в спинную аорту, которая идет под хордой к хвостовому отделу зародыша. Дойдя до конца тела, они делает петлю и переходит в хвостовую вену, идущую к голове. В районе анального отверстия хвостовая вена переходит в заднюю кардинальную вену. Здесь от кардинальной вены отходят по четыре сосуда с каждой стороны кишки, снабжающие кровью подкишечную вену. В передней части тела зародыша задняя кардинальная вена сливается с передней кардинальной веной и впадает в венозный синус сердца. Подкишечная вена впадает в печень, расположенную в задней части желточного мешка. Из печени кровь по печеночной вене выходит на правую сторону желточного мешка, охватывая его нижнюю часть, и снизу переходит на левую сторону. Эта система сосудов носит название печеночно-желточной вены и выполняет дыхательную функцию (Крыжановский, 1933; Соин, 1968). На левой стороне желточного мешка капилляры сливаются в собирательный сосуд печеночно-желточной вены, который, поднимаясь вверх, впадает в венозный синус сердца.
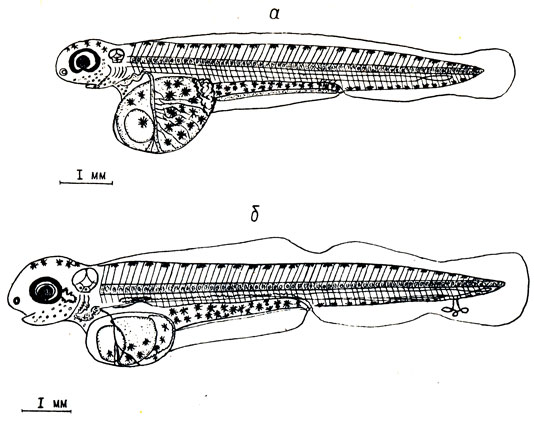
Рис. 4. Седьмой и восьмой этапы эмбрионального развития пыжьяна: а - возраст 74 суток, наибольшее развитие кровеносной системы на желточном мешке. Оболочка снята, б - возраст 102 суток, зародыш пород выклевом. Оболочка снята
Голова снабжается кровью через сонные артерии, которые образуют здесь сложную систему капилляров. Они сливаются в передние кардинальные вены, по которым кровь попадает в венозный синус сердца.
Седьмой этап характеризуется появлением жаберного кровообращения, обособлением челюстей, ростом жаберных крышек, образованием роговицы, заканчивается формированием на жаберных дугах жаберных лепестков. При средней температуре воды 2,2° этап длится с 62-х по 87-е сутки.
В возрасте 64 суток (сумма тепла 163,2°) зародыш под оболочкой подвижен, при раздражении переворачивается. Длина его составляет 8,2 мм, длина кишки - 2,8 мм. На нижней части головы появились железы вылупления. Кровь из сердца в спинную аорту поступает по челюстным, а также по четырем жаберным дугам аорты.
В возрасте 74 суток (сумма тепла 186,5°) длина зародыша равна 9,0 мм. Вынутые из оболочки зародыши плавают в течение нескольких минут. Жаберная крышка еще не доходит до жаберных дуг, ее длина равна 0,35 мм. Появляется зачаток нижней челюсти. Железы вылупления становятся многочисленнее и располагаются на нижней части головы, вокруг глаз и на перикарде (рис. 4, а).
Желточный мешок продолжает редуцироваться, его размеры составляют 1,8×1,5 мм. Диаметр жировой капли равен 1,25 мм. Кровеносная система на желточном мешке сильно развита. На правой стороне мешка капилляры печеночно-желточной вены занимают почти всю поверхность. На левой стороне они охватывают снизу большую часть поверхности. Собирательный сосуд печеночно-желточной вены проходит по границе с жировой каплей и впадает в венозный синус сердца.
На 78-е сутки развития начинается дифференцировка плавниковой каймы на спинной и жировой плавники. Происходит дальнейшее развитие жаберной крышки - они прикрывает одну жаберную дугу. Нижняя челюсть также увеличивается в размерах и достигает вертикали, проходящей через заднюю треть глаза. Размеры глазного бокала и слуховой капсулы составляют соответственно 0,70×0,60 мм и 0,62×0,45 мм. Желточный мешок продолжает редуцироватья (размеры 1,6×1,3 мм). Пигментация особенно сильно развита вдоль кишки, на желточном мешке и концах миотомов.
В кровеносной системе происходит ряд изменений. От спинной аорты отходят передняя и задняя брыжеечные артерии, снабжающие кровью кишку. Появляются первые сегментные сосуды, отходящие от спинной аорты в передней части туловищного и в хвостовом отделе. Сегментные сосуды идут вдоль передней миосепты вверх, делают петлю, и, спускаясь по заднему краю миотома, впадают в заднюю кардинальную вену.
В возрасте 87 суток (сумма тепла 215,0°) длина зародыша составляет 9,0 мм. Находясь в оболочке, зародыш периодически переворачивается. При снятии оболочки длительное время плавает. Жаберная крышка, прикрывающая первую и половину второй жаберной дуги, и нижняя челюсть, достигающая вертикали, проходящей через середину глаза, приобретают подвижность. В глазном бокале образуется роговица. Железы вылупления становятся более многочисленными.
По-прежнему достаточно хорошо развита дыхательная сеть сосудов на желточном мешке. Сегментные сосуды появляются во всех сегментах тела. В хвостовом отделе происходит редукция меланофоров на месте будущей хвостовой жабры.
Размеры желточного мешка уменьшаются и составляют 1,45×0,85 мм. Впереди жировой капли виден остаток перибластического синуса.
Заключительный восьмой этап развития начинается с появления первых жаберных лепестков на жаберных дугах и заканчивается выклевом личинок. При средней температуре воды 4,4° этап продолжается с 88-х по 135-е сутки.
На 88-е сутки развития (сумма тепла 217,4°) на жаберных дугах появляются лепестки: на третьей дуге четыре, на четвертой - три. Над желточным мешком имеется зачаток почки. У зародышей уменьшается ток крови в подкишечной вене. Дыхательная сеть сосудов на желточном мешке начинает редуцироваться. Основной ток крови осуществляется по задней кардинальной вене.
В возрасте 95 суток (сумма тепла 233,6°) длина зародыша составляет 10,0 мм. Вынутый из оболочки зародыш теперь жизнеспособен. Жаберные крышки увеличиваются в размере и прикрывают две передние жаберные дуги. Глазные бокалы приобрели подвижность.
В кровеносной системе произошли следующие изменения: уменьшился ток крови по подкишечной вене и по капиллярным сосудам печеночно-желточной вены. В качестве компенсации уменьшения дыхательной поверхности желточного мешка происходят изменения в строении хвостовой вены. Она разветвляется на 2 - 3 сосуда и выходит в нижнюю часть лопасти хвостового плавника в виде Т-образного сосуда - хвостовой жабры.
У зародышей в возрасте 102 суток хвостовая жабра имеет сложное строение, а позади глаз появляются псевдобранхии (см. рис. 4, б).
В возрасте 117 суток (сумма тепла 314,8°) зародыш имеет длину 10,2 мм, размеры желтка - 0,45×0,57 мм. Наиболее интенсивно пигментированы кишечник и нижняя половина желточного мешка. Имеются также меланофоры на верхней части головы и на концах миотомов. Сформировались зачатки спинного и жирового плавников. У зародышей сохраняется разветвленная хвостовая вена и хвостовая жабра. Псевдобранхии приобретают сложное строение. Функционируют сегментные сосуды. Жаберная крышка закрывает первую, вторую и половину третьей жаберной дуги. Образовались расширенный желудок и более узкий пищевод. В кишечнике появилась складчатость, и он получил способность к перистальтическим движениям. В его полости обнаруживаются остатки полупереваренного желтка, перемещающиеся к анальному отверстию. На этой стадии наблюдается начало выклева.
В возрасте 135 суток (сумма тепла 424,1°) выклев заканчивается. Выклюнувшиеся личинки имеют длину 10,3 мм; размеры желточного мешка составляют 1,00×0,65 мм, желтка - 0,40×0,51 мм. Пигментация тела становится менее интенсивной. Жаберная крышка закрывает две-три жаберных дуги. Длина грудных плавников составляет 1,6 мм. У личинок имеются псевдобранхии, хвостовая жабра у некоторых личинок редуцируется.
На этом заканчивается эмбриональный период развития сига-пыжьяна.
Обсуждение результатов и выводы. Эмбриональное развитие пыжьяна происходит по общему для сиговых типу (Кузьмин, 1963; Смольянов, 1966; Буланов, 1976; Лебедева, 1982; Черняев, 1968, 1982; Кугаевская, Нечаева, 1984). По времени наступления и длительности этапов развития пыжьян более близок к сигу-лудоге и чудскому сигу (Кубрак, 1960; Маненкова, 1972; Лебедева, 1981). Однако длительность инкубации икры пыжьяна на ЦЭС "Ропша" гораздо меньше, что обусловлено более высокой температурой воды, особенно на ранних стадиях эмбрионального развития. Среднемесячная температура воды во время инкубации колебалась от 2,2 до 5,2°. При сравнении средней температуры воды на ЦЭС "Ропша" и Тобольском рыбоводном заводе, где уже в течение ряда лет успешно проходит инкубирование икры обского пыжьяна (отход за время инкубации не превышает 15%), выясняется, что температура воды на ЦЭС "Ропша" в отдельные месяцы выше в 2 - 10 раз. В 1982 - 1983 гг. эмбриональное развитие пыжьяна на ЦЭС "Ропша" продолжалось 135 суток (сумма тепла 424,1°), на Тобольском рыбозаводе - 207 суток (сумма тепла 155,8°). Уменьшение длительности инкубации влечет за собой ранний выклев личинок. При посадке их в водоемы кормовая база еще недостаточно развита, что обусловливает снижение темпа роста и выживаемости сеголеток.
Кроме того, высокая температура воды во время инкубации икры ведет к снижению выхода личинок и возрастанию числа неправильно развивающихся эмбрионов. Так, по литературным данным (Colby, Brooke, 1970), повышение температуры воды при инкубации икры Goregonus clupeaformis (вида, близкого к Coregonus lavaretus pidschian) от оптимальной величины 0,5° до 7,8° приводит к снижению выхода личинок с 72,7 до 18,8%, увеличению числа уродливых эмбрионов до 25% и уменьшению размеров выклюнувшихся личинок.
При инкубации икры пыжьяна на ЦЭС "Ропша" в условиях повышенной температуры воды выход личинок снижается до 30%. Средняя длина выклюнувшихся личинок составляет 10,3 мм. Эти показатели ниже, чем при инкубации икры других сиговых рыб в более благоприятных условиях (Юхнева, 1967; Кугаевская, Нечаева, 1984, и др.). Отсюда вытекает необходимость использования воды с более низкой температурой при инкубировании икры сига пыжьяна.
Буланов Д. П. 1976. Эмбриональное развитие кубенской нельмы. - Изв. ГосНИОРХ, 118.
Крыжановский С. Г. 1933. Органы дыхания личинок рыб и псевдобранхия.- Труды лаб. эволюционной морфологии АН СССР, 1, 2.
Крыжановский С. Г., Дислер Н. Я., Смирнова Е. Н. 1953. Эколого-морфологические закономерности развития окуневидных рыб. - Труды ИЭМЭЖ им. Северцова АН СССР, 10.
Кубрак И. Ф. 1960. Эмбриональное развитие чудского сига, акклиматизируемого в прудах Молдавии. - Труды ин-та биологии Молд. фил. АН СССР, 2, 2.
Кугаевская Л. В., Нечаева Н. Л. 1984. Работа с икрой сиговых на Тобольском рыбоводном заводе. - Рыбное хоз-во, 12.
Кузьмин А. Н. 1963. Эмбриональное развитие пеляди. - Труды Обь-Та-зовск. отд.-ния ГосНИОРХ, 3.
Лебедева О. А. 1981. Развитие икры и личинок чудского сига Coregonus lavaretus maraenoides (Poljakow) - Сб. научн. трудов ГосНИОРХ, 167.
Лебедева О. А. 1982. Эмбрионально-личиночное развитие чира Coregonus nasus (Pall.) и муксуна Coregonus muksun (Pall.). - Сб. научн. трудов ГосНИОРХ, 185.
Маненкова Г. М. 1972. Эмбриональное и раннее постэмбриональное развитие ладожского сига-лудоги.- В сб.: Краткие тез. докл. к IV совещ. молодых научн. работников ГосНИОРХ. Л. Изд. ГосНИОРХ.
Руденко Г. П. 1975. Характеристика малых озер и перспективы их рыбохозяйственного использования. - Изв. ГосНИОРХ, 99.
Руденко Г. П. 1978. Биологические основы товарного выращивания поликультуры ценных видов рыб.- Изв. Псковск. отд-ния ГосНИОРХ, 3.
Смольянов И. И. 1957. Развитие белорыбицы, нельмы и сига-нельмушки. - Труды ИЭМЭЖ им. Северцова АН СССР, 20.
Смольянов И. И. 1966. Эмбриональное развитие муксуна Coregonus muksun (Pall.) - Вопр. ихтиологии, 6, 1 (38).
Соин С. Г. 1968. Приспособительные особенности развития рыб. М.
Черняев Ж. А. 1968. Эмбриональное развитие байкальского омуля. М.
Черняев Ж. А. 1982. Воспроизводство байкальского омуля. М.
Юхнева В. С. 1967. Наблюдение за нерестом и развитием икры сиговых рыб па р. Сыня.- В кн.: Озерное и прудовое хозяйство в Сибири и на Урале. Тюмень.
Colby P. J., Brooke L. T., 1970. Survival and development of lake herring (Coregonus artedii) eggs at various incubation temperatures. - In: Biology of Coregonid fishes. Winnipeg.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'