Роль сфериид в продукции макробентоса мезотрофного озера Калищевского Лужского района (В. Ф. Шуйский)
УДК 574.587
Весьма актуальной и сложной задачей современной гидробиологии является изучение особенностей продуцирования бентоса в условиях антропического воздействия на экосистемы континентальных водоемов. Решению подобной задачи должно предшествовать детальное исследование продукционного процесса донных сообществ и их компонентов в неудобряемых озерах. Значительную и многогранную роль в жизни пресноводных экосистем играют двустворчатые моллюски семейства Sphaeriidae. Они входят в рацион рыб-бентофагов, способствуют процессу самоочищения водоемов, принимают участие в образовании паразитарных связей (Алимов, 1981). Целью настоящей работы являлось выяснение роли сфериид в продукции макробентоса одного из мезотрофных озер Лужского района, где сосредоточено большое количество экспериментальных удобряемых водоемов ГосНИОРХ. Полученные нами данные могут послужить исходной основой при сравнительном анализе влияния удобрений на озера.
Озеро Калищевское относится к бассейну р. Оредеж и имеет речное происхождение. Выток из озера незначителен, осуществляется через небольшую протоку. Питание происходит за счет поверхностного стока и осадков. Площадь дна озерной котловины - 33 га, максимальная глубина - 11,5 м, средняя - 6,8 м, объем водной массы - 2,2 млн.м2., индекс развития береговой линии - 2,4. По нашим данным, литораль занимает 8,1% площади (4,2% - илистый песок, 3,9% - слабозаиленный песок), сублитораль - 9,0% (ил с детритом), профундаль - 82,9% (78,8% - бурый слабоминерализованный ил, 2,3% - черный гомогенный маслянистый ил, 1,8% - белый кварцевый песок, белая глина). Площадь зарастания озера составляет 10%, преобладает растительность с плавающими листьями -94% зарастающей площади (по устному сообщению С. В. Мещеряковой).
Отбор проб осуществлялся ежедекадно на 9 станциях в течение лета 1982 г. (с 5.06 по 27.08). Кроме того, в конце июля за короткий срок (10 суток) была отобрана 71 проба в различных участках акватории. Орудиями сбора служили дночерпатель системы Петерсена и стратометр С-1, для промывания проб использовалось газовое сито № 37.
По данным о структуре грунтов на этой 71 станции выделили характерные типы субстрата и определили занимаемую ими площадь. Величины биомассы бентоса на разных биотопах нашли по данным 71 пробы (BII) и одновременно по материалам основных станций (BI). Величины BII/BI, отражающие репрезентативность каждой из станций, использовались при расчете биомассы и продукции бентоса на соответствующих биотопах: Bt=B't (BII/BI)·S; Pt=P't (BII/BI)·S, где Bt (Pt) - средняя биомасса (продукция) донной фауны биотопа ко времени t, B't (P't) - биомасса (продукция) бентоса на основной станции, S (м2.) - площадь данного биотопа. При анализе динамики численности моллюсков использовался метод "средней взвешенной" для устранения флуктуации на тренде.
Для расчета продукции сфериид физиологическим способом определили параметры уравнения обмена Sphaerium corneum. Измерение потребленного кислорода проводилось методом замкнутых сосудов при температуре 20°С (период адаптации - не менее суток). Масса моллюсков варьировала от 8,0·10-4 до 1,2 г. Содержание кислорода определяли по Винклеру. После обсушивания моллюсков взвешивали (вместе с раковиной и водой в мантийной полости), затем измеряли. Параметры степенных зависимостей массы от длины раковины и скорости дыхания от массы моллюсков рассчитывали методом наименьших квадратов (Сущеня, 1972; Умнов, 1976).
Продукцию доминирующих видов донных животных (встречаемость в пробах больше 0,6, биомасса - не менее 10% от общей биомассы бентоса) определяли физиологическим способом. Расчет продукции остальных гидробионтов производили с помощью имеющихся в литературе P/B-коэффициентов и уравнений вида dW/dt = aWb (Гаврилов, 1970; Тодераш, 1979; Панкратова, Балушкина, 1981; Голубков, 1982). Рацион хищных беспозвоночных (C), необходимый для определения суммарной продукции бентоса, рассчитывали по балансовому равенству C = P + T + U (P - прирост, T - траты на обмен, U - неусвоенная часть пищи). Долю усвояемой ими пищи [(P + T)/C·100%] приравнивали к 80% (Винберг, 1970); долю животной пищи в рационе считали равной 70% (Гаврилов, 1970).
Калорийность моллюсков считали равной 330 кал/г, K2 = 0,26 (Алимов, 1981). Величину суточной продукции моллюсков как функцию обмена находили по уравнению: Pt = nPi; Pi = 1/ [Q10(20° - t°)/10]· R (мгO2/час)·24 (час) · 3,4 (кал/мгO2) · (1/330) (г/кал) · [0,26/ (1 - 0,26)] = Q100,1 (t° - 20°) · 0,0869 · a · Wb (г); R (мгO2/час) = aWb (г), где Pt (г) - продукция сфериид за сутки, n (экз.) - численность сфериид к моменту t, Pt (г) - продукция за сутки особи со средней массой (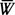 ), R -скорость потребления кислорода моллюсками с массой W. Продукцию сфериид за летний период находили интегрированием (по формуле трапеций). Для контроля продукцию сфериид на литорали за интервал времени 10.07 - 9.08.83 г. (минимальное отмирание моллюсков, завершивших отрождение) рассчитали также с помощью метода, основанного на измерении элиминации моллюсков (Шуйский, 1984).
), R -скорость потребления кислорода моллюсками с массой W. Продукцию сфериид за летний период находили интегрированием (по формуле трапеций). Для контроля продукцию сфериид на литорали за интервал времени 10.07 - 9.08.83 г. (минимальное отмирание моллюсков, завершивших отрождение) рассчитали также с помощью метода, основанного на измерении элиминации моллюсков (Шуйский, 1984).
Рис. 1. Динамика численности (1 - N, экз/м2.), средней массы особи (2 -
, мг) и биомассы (3 - В, г/м2.) сфериид на литорали (а - илистый песок, б - слабозаиленный песок) и сублиторали (в) оз. Калищевского по материалам основных станций летом 1982 г.Распределение сфериид по биотопам оказалось достаточно видоспецифично. Шаровки (Sphaerium corneum, S. scaldianum) распространены исключительно на литорали. Горошинки населяют как литораль (Pisidium pulchellum, P. subtruneatum), так и сублитораль (P. supinum). Виды P. obtusalis и P. ponderosum малочисленны и встречаются в основном на небольших глубинах1. Величины численности и биомассы сфериид значительно варьируют на различных грунтах литорали. Повышенное заиление субстрата ведет к снижению численности сфериид, что отмечалось многими авторами (Саватеева, 1964; Алимов, 1969, и др.). Роль сфериид в структуре локальных донных сообществ оз. Калищевского весьма различна. Так, доля их в составе биомассы бентоса литорали и сублиторали колебалась от 0 до 93%.
1 (Выражаю искреннюю признательность докт. биол. наук, ст. науч. сотруднику ЗИН АН СССР А. Ф. Алимову за помощь, оказанную при определении моллюсков)
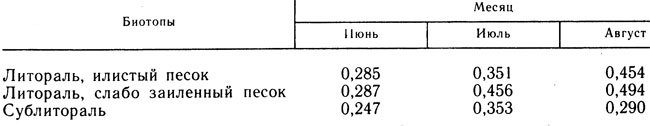
Таблица 1. Значения месячных P/B-коэффициентов сфериид для летнего периода на различных биотопах оз. Калищевского (1982 г.)
Основными показателями, характеризующими состояние популяций сфериид и необходимыми при расчете продукции физиологическим методом, являются биомасса, численность и средняя масса особи. Динамика данных показателей на разных биотопах за летний период отражена на рис. 1. Следует отметить, что отрождение молоди у сфериид было значительно растянуто во времени (60 - 65 суток) из-за неодновременного достижения ими половозрелости (Алимов, 1967) и неоднородного видового состава. Кроме того, существенное влияние на процесс вымета молоди оказывала динамика температуры. Так, похолодание в июне вызвало задержку отрождения (аналогичное явление описал И. И. Десятик, 1968); с уменьшением температуры к началу осени отрождение прекратилось. С другой стороны, в июле и августе периоды потепления сопровождались интенсификацией отрождения. Динамика состояния популяций имела более плавный характер на сублиторали, чем на литорали, где колебания температуры значительней, а видовой состав богаче. Отмирание основной части отметавших особей происходит в августе; к началу осени элиминация моллюсков уже на 93,6% обусловлена выеданием.
Наиболее широко распространен на литорали оз. Калищевского моллюск S. corneum. Поэтому для расчета продукции сфериид были экспериментально определены параметры уравнения зависимости скорости потребления S. corneum кислорода (R, мгO2/час) от его массы (W, в граммах сырой массы): R = (2,6·10-2±1,53·10-2·W(6,52·10-1±8,21·10-2). Средняя масса использованных в опытах моллюсков была 5,4·10-3, коэффициент корреляции r = 0,9430. Продукция рассчитывалась и с использованием другого уравнения обмена сфериид (Алимов, 1981): R = (1,29·10-1±5,7·10-2)·W(8,95·10-1). Очевидно, что уровень обмена S. Corneum в оз. Калищевском сравнительно низок (параметры приведенных уравнений достоверно различаются при P = 90%). Анализ возможных причин наблюдаемых различий проведен нами ранее (Шуйский, 1983). Продукция сфериид на литорали за период 10.07 - 9.08.83 г., определенная с помощью данных уравнений, равна соответственно 0,486 и 0,606 г/м2. При использовании метода расчета продукции, основанного на измерении элиминации, данная величина не превышает 0,430 г/м2., поэтому для дальнейших расчетов применялись параметры уравнения обмена, найденные экспериментально.
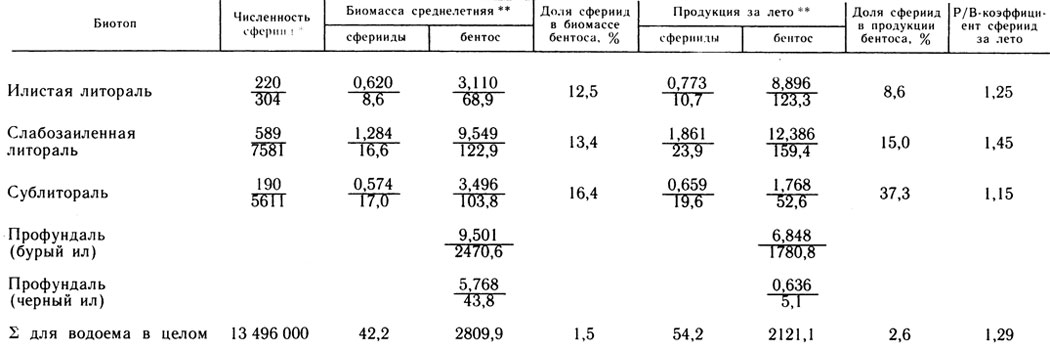
Таблица 2. Численность, биомасса и продукция сфериид и всего бентоса на различных биотопах оз. Калищевского (1982 г.)
В табл. 1 приведены P/B-коэффициенты сфериид различных биотопов для каждого летнего месяца. Очевидно, что продукционный процесс в течение изученного периода протекает неравномерно. Наибольшие значения коэффициентов для литорали приходятся на июль и август, в условиях сублиторали месячный P/B-коэффициент достигает максимальной величины в июле. Это отражает характерные для разных зон различия в сроках отрождения моллюсками основной части молоди. Определено значение летнего (за 3 месяца) P/B-коэффициента сфериид оз. Калищевского, усредненное по всем биотопам: P/B = 1,29±0,091. Полученное значение коэффициента вполне сопоставимо с величинами, приводимыми в литературе для Pisidium: 0,80 - 2,16 (Алимов, 1981), 1,0 - 1,5 (Holopainen, 1979) и для Sphaerium: 1,44 - 1,53 (Арабина, 1966), 1,5 (Арабина, 1968). Рассчитаны также летние P/B-коэффициенты других доминирующих видов: Sialis lutaria (3,5), Einlcldia ex gr. carbonaria (3,4), Procladius ferrugineus (4,4), Chironomus plumosus (0,7). Найденные P/B-коэффициенты хирономид близки к аналогичным величинам, полученным для указанных видов на мезотрофном озере Мястро (Винберг и др., 1971).
Значения биомассы, продукции сфериид и всего бентоса и данные о соотношении этих величин для разных биотопов приведены в табл. 2. Очевидно, что роль сфериид в общем продукционном процессе бентоса водоема во многом определяется соотношением площади дна, занимаемой литоралью и сублиторалью, и площади профундали. Биомасса моллюсков семейства Sphaeriidae в оз. Калищевском не превышала 1,5% от общей биомассы донных животных. Продукция сфериид за лето достигала 2,6% от продукции бентоса. Это обусловлено тем, что сферииды, предпочитая небольшие глубины, обитают в хорошо прогреваемых грунтах. В условиях профундали, наоборот, преобладают низкие летние температуры (10 - 12°), замедляющие процесс продуцирования донных животных.
Величины продукции бентосного сообщества в значительной степени обусловлены степенью развития хищных беспозвоночных и, следовательно, величиной их рациона (рис. 2). Существенно, что сферииды, защищенные раковиной, не входят в состав рациона хищного бентоса. Вследствие этого доля моллюсков семейства Sphaeriidae в продукции бентоса литорали примерно равна их доле в биомассе, несмотря на сравнительно невысокую скорость продуцирования. На сублиторали, где особенно развит пресс хищных беспозвоночных, за счет сфериид образуется 16% биомассы и 37% (т. е. в 2,3 раза больше) продукции бентоса.
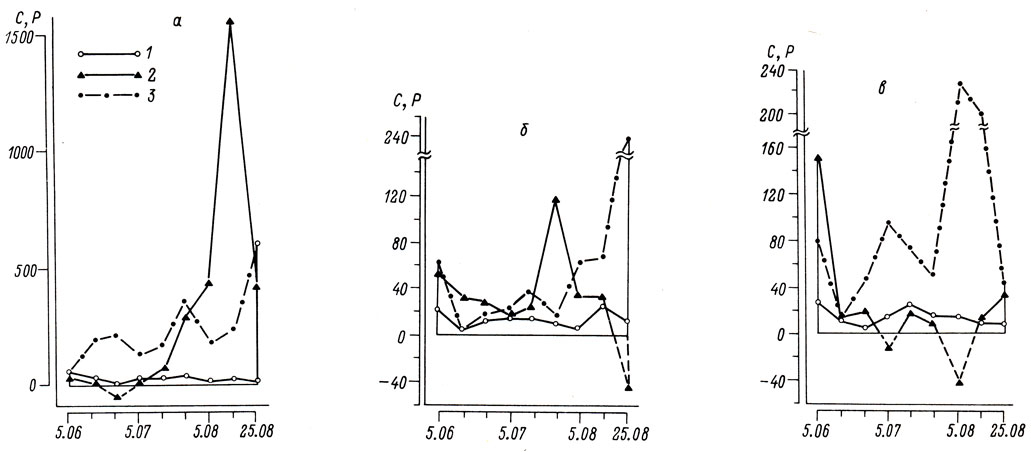
Рис. 2. Ход изменения величин суточной продукции сфериид (1 - P, мг/м2 и всего макробентоса (2) и рациона хищных беспозвоночных (3 - C, мг/м2) в условиях литорали (а - илистый песок, б - слабозаиленный песок) и сублиторали (в) оз. Калищевского летом 1982 г.
Итак, сферииды распределены по биотопам видоспецифично и крайне неравномерно, что объясняется особенностями условий обитания на различных глубинах и грунтах. Отрождение молоди у сфериид растянуто во времени (около 60 суток). Этому способствуют неоднородность видового состава и неодновременность достижения половозрелости. Отрождение и отмирание сфериид заканчивается к началу осени. Динамика температуры значительно влияет на интенсивность отрождения. Благоприятный температурный режим грунтов мелководий способствует ускорению продукционного процесса в популяциях сфериид. Летний (за 3 месяца) P/B-коэффициент моллюсков семейства Sphaeriidae для оз. Калищевского равен 1,29±0,091. Специальные опыты по изучению элиминации моллюсков показали, что сферииды активно выедаются рыбами-бентофагами (Шуйский, 1984). Таким образом, сферииды могут играть весьма важную роль в продукции макробентоса озер с выраженными зонами литорали и сублиторали. При интенсивном развитии хищного бентоса эта роль существенно увеличивается.
Алимов Л. Ф. 1967. Особенности жизненного цикла и роста пресноводного моллюска Sphaerium corneum (L.) - Зоол. журн., 46, 2.
Алимов А. Ф. 1969. Некоторые сведения по фауне и экологии сфериид Калининградской области. - В кн.: Гидробиол. исследования, 5. Таллин.
Алимов А. Ф. 1981, Функциональная экология пресноводных двустворчатых моллюсков. Л.
Арабина И. П. 1966. Рост пресноводного моллюска Sphaerium corneum. - ДАН БССР, 10, 12.
Арабина И. П. 1968. Итоги изучения зообентоса промысловых озер Нарочанской группы. - В сб.: Третья зоол. конф. БССР. Тез. докл. Минск.
Винберг Г. Г. 1970. Общие особенности экологической системы оз. Дривяты. - Труды ВГБО, 13.
Винберг Г. Г., Бабицкий В. А., Гаврилов С. И. и др. 1971. Биологическая продуктивность озер разного типа. - В кн.: Биопродуктивность озер Белоруссии. Минск.
Гаврилов С. И. 1970. Бентос озера Дривяты и его продуктивность. - Труды ВГБО, 15.
Голубков С. М. 1982. Соотношение скоростей роста и энергетического обмена у пресноводных беспозвоночных. Автореф. канд. дис. Минск.
Десятик И. И. 1968. Соотношение веса и линейных размеров у некоторых видов пресноводных моллюсков. - ДАН БССР, 12, 9.
Панкратова В. Я., Балушкина Е. В. 1981. Зависимость массы тела от длины и интенсивности обмена от массы тела у личинок хирономид. - В кн.: Основы изучения пресноводных экосистем. Л.
Саватеева Е. Б. 1964. О распространении и экологии моллюсков семейства Sphaeriidae в некоторых озерах Карелии. - Изв. ВНИОРХ, 57.
Сущеня Л. М. 1972. Интенсивность дыхания ракообразных. Киев.
Тодераш И. К. 1979. Энергетический баланс личинок хирономид. - В кн.: Общие основы изучения водных экосистем. Л.
Умное А. А. 1976. Применение статистических методов для оценки параметров эмпирических уравнений, описывающих взаимосвязь между энергетическим обменом и массой тела животного. - Журн. общ. биологии, 37, 1.
Шуйский В. Ф. 1983. Некоторые аспекты биологии пресноводного двустворчатого моллюска Sphaerium corneum в мезотрофном озере.- Сб. науч. трудов ГосНИОРХ, 204.
Шуйский В. Ф. 1984. Расчет продукции сфериид в условиях интенсивного воздействия на них пресса рыб-бентофагов (на примере мезотрофного озера Калищевского Лужского района). - В кн.: Проблемы рыбохозяйственных исследований внутренних водоемов Северо-Запада европейской части СССР. Петрозаводск.
Holopainen I. J. 1979. Population dynamiCs and production of Pisidium species (Bivalvia, Sphaeriidae) in the oligotrophic and mesotrophic lake Paajarvi, Southern Finland. - Arch. Hydrobiol, 54.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://ribovodstvo.com/ 'Рыбоводство'